

Targeted overexpression of the neurite growth-associated protein B-50/GAP-43 in cerebellar Purkinje cells induces sprouting after axotomy but not axon regeneration into growth-permissive transplants
- PMID: 9348347
- PMCID: PMC6573080
- DOI: 10.1523/JNEUROSCI.17-22-08778.1997
Targeted overexpression of the neurite growth-associated protein B-50/GAP-43 in cerebellar Purkinje cells induces sprouting after axotomy but not axon regeneration into growth-permissive transplants
Abstract
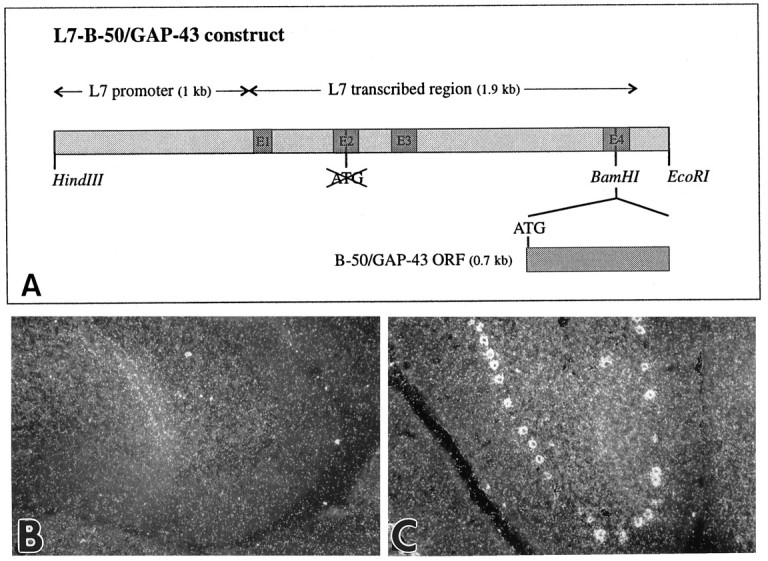
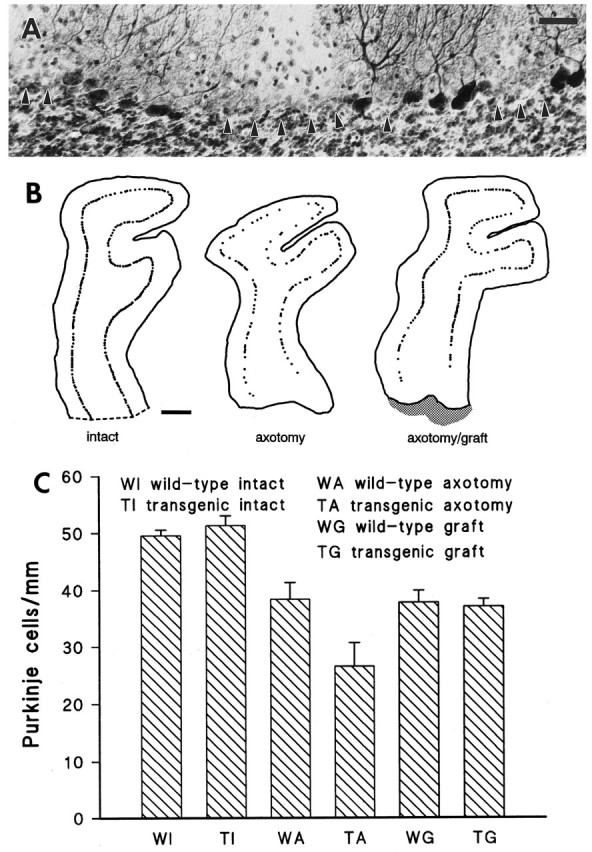
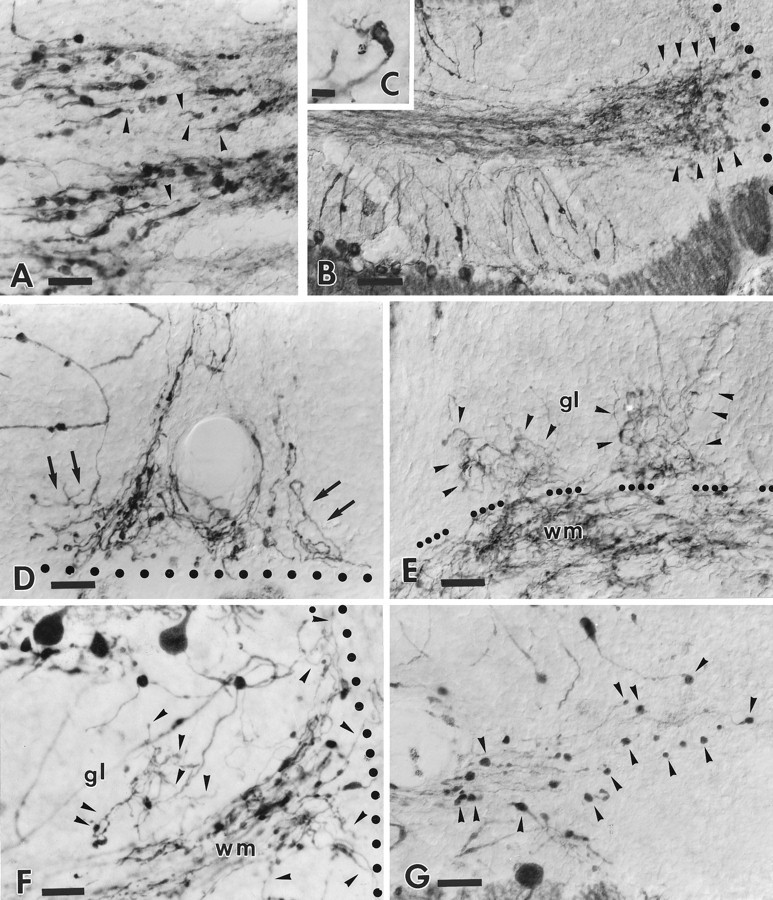
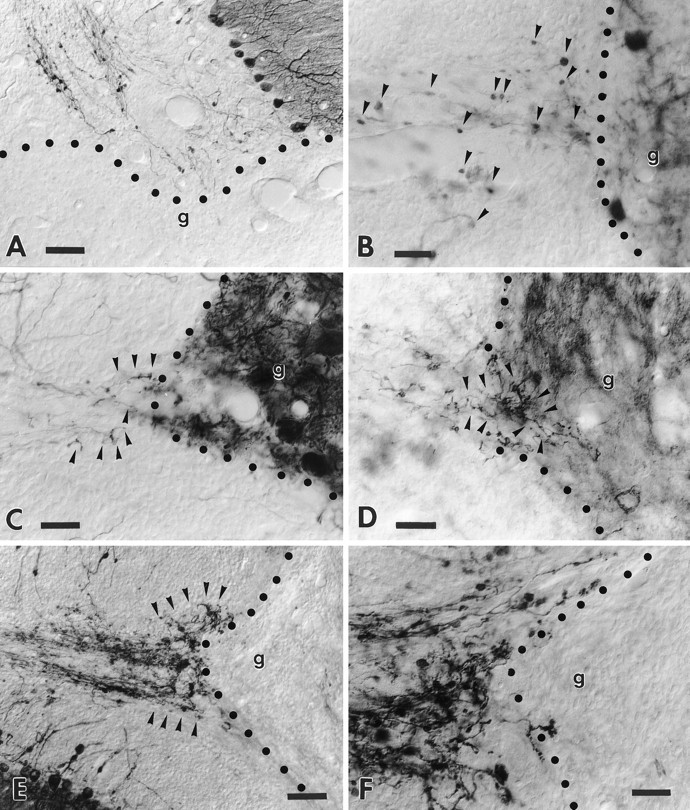
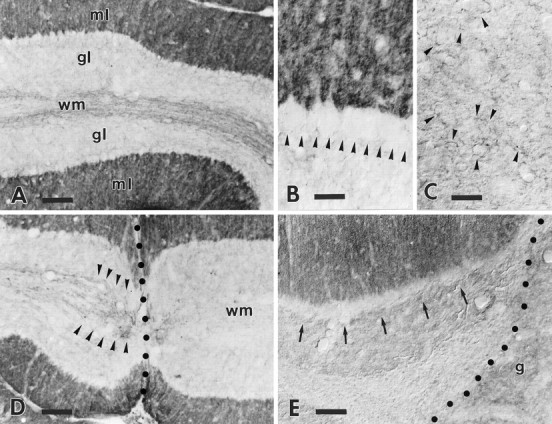
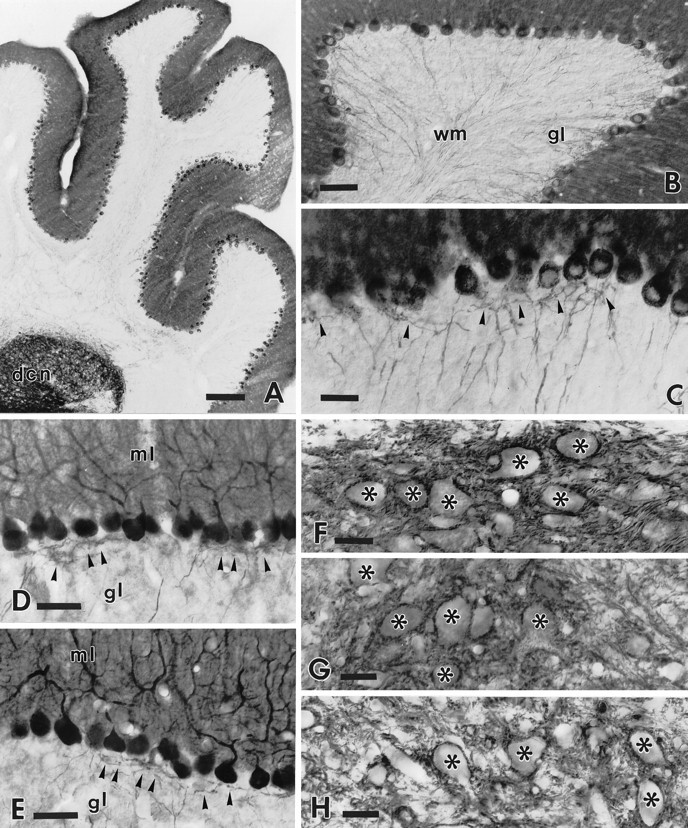
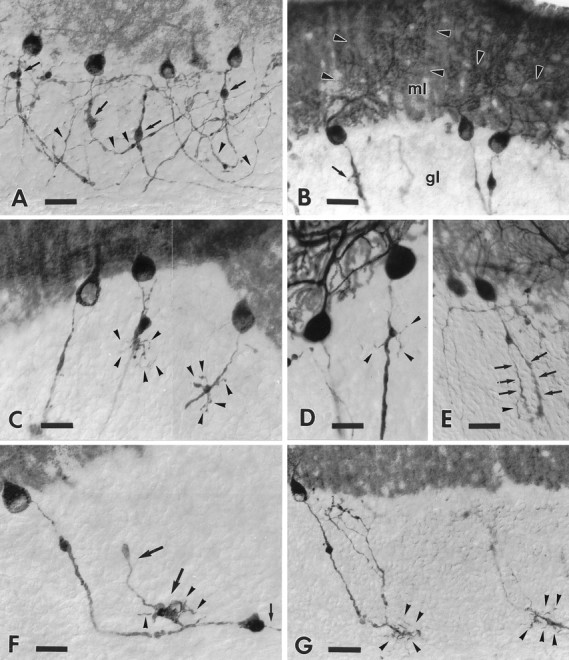
B-50/GAP-43 is a nervous tissue-specific protein, the expression of which is associated with axon growth and regeneration. Its overexpression in transgenic mice produces spontaneous axonal sprouting and enhances induced remodeling in several neuron populations (; ). We examined the capacity of this protein to increase the regenerative potential of injured adult central axons, by inducing targeted B-50/GAP-43 overexpression in Purkinje cells, which normally show poor regenerative capabilities. Thus, transgenic mice were produced in which B-50/GAP-43 overexpression was driven by the Purkinje cell-specific L7 promoter. Uninjured transgenic Purkinje cells displayed normal morphology, indicating that transgene expression does not modify the normal phenotype of these neurons. By contrast, after axotomy numerous transgenic Purkinje cells exhibited profuse sprouting along the axon and at its severed end. Nevertheless, despite these growth phenomena, which never occurred in wild-type mice, the severed transgenic axons were not able to regenerate, either spontaneously or into embryonic neural or Schwann cell grafts placed into the lesion site. Finally, although only a moderate Purkinje cell loss occurred in wild-type cerebella after axotomy, a considerable number of injured transgenic neurons degenerated, but they could be partially rescued by the different transplants placed into the lesion site. Thus, B-50/GAP-43 overexpression substantially modifies Purkinje cell response to axotomy, by inducing growth processes and decreasing their resistance to injury. However, the presence of this protein is not sufficient to enable these neurons to accomplish a full program of axon regeneration.
Figures
References
-
- Aigner L, Arber S, Kapfhammer JP, Laux T, Schneider C, Botteri F, Brenner H-R, Caroni P. Overexpression of the neural growth-associated protein GAP-43 induces nerve sprouting in the adult nervous system of transgenic mice. Cell. 1995;83:269–278. - PubMed
-
- Barron KD. Neuronal responses to axotomy: consequences and possibilities for rescue from permanent atrophy or cell death. In: Seil FJ, editor. Neural regeneration and transplantation. Alan Liss; New York: 1989. pp. 79–99.
-
- Baürle J, Grover BG, Grüsser-Cornehls U. Plasticity of GABAergic terminals in Deiters nucleus of Weaver mutant and normal mice: a quantitative light microscopic study. Brain Res. 1992;591:305–318. - PubMed
-
- Baürle J, Oestreicher AB, Gispen WH, Grüsser-Cornehls U. Lesion-specific pattern of immunocytochemical distribution of growth associated protein B-50 (GAP-43) in the cerebellum of weaver and PCD-mutant mice: lack of B-50 involvement in neuroplasticity of Purkinje cell terminals? J Neurosci Res. 1994;38:327–335. - PubMed
-
- Benowitz LI, Routtenberg A. GAP-43: an intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 1997;20:84–91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous