

The olivocerebellar projection mediates ibogaine-induced degeneration of Purkinje cells: a model of indirect, trans-synaptic excitotoxicity
- PMID: 9348351
- PMCID: PMC6573067
- DOI: 10.1523/JNEUROSCI.17-22-08828.1997
The olivocerebellar projection mediates ibogaine-induced degeneration of Purkinje cells: a model of indirect, trans-synaptic excitotoxicity
Abstract
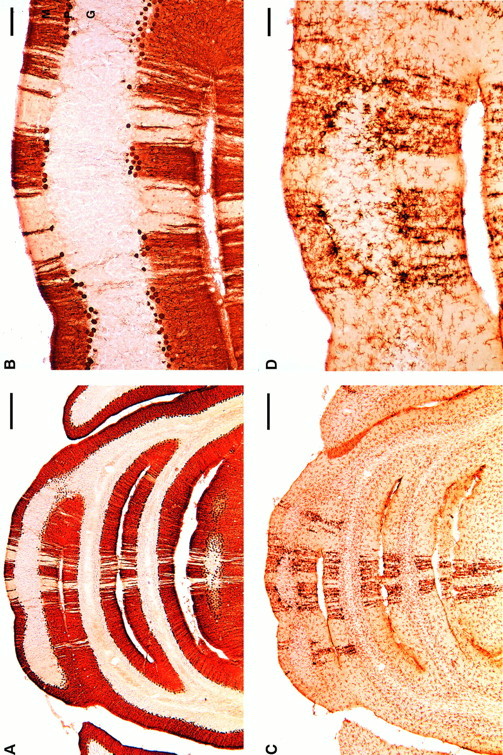
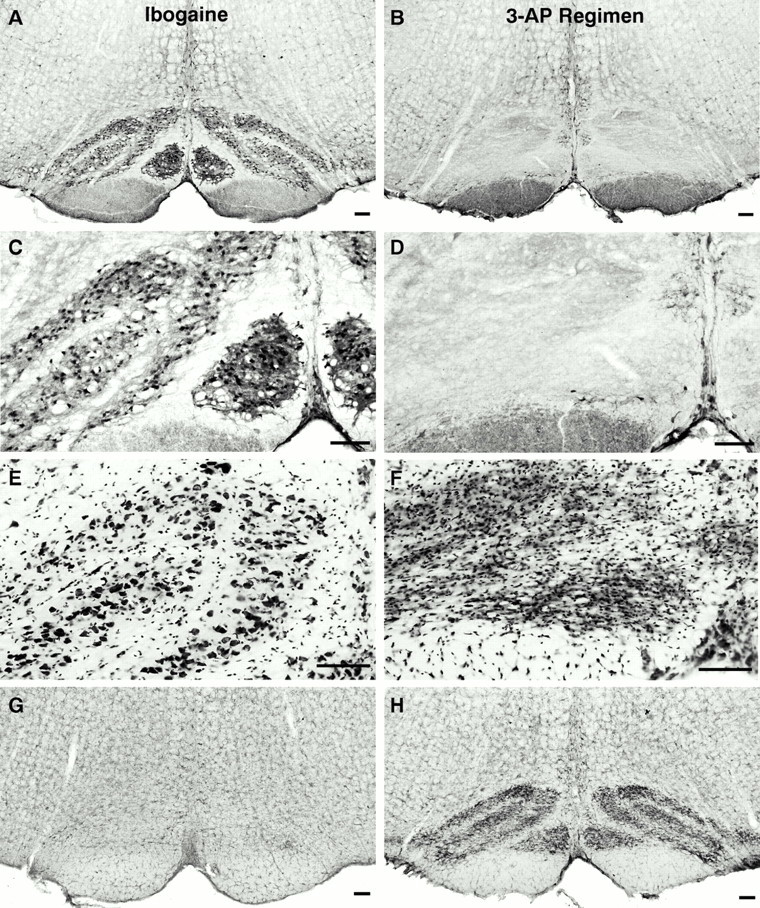
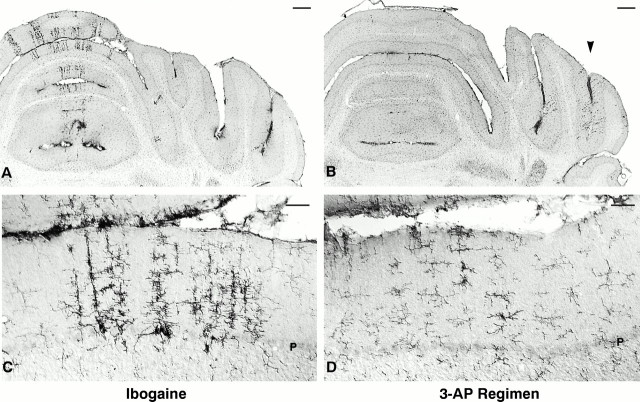
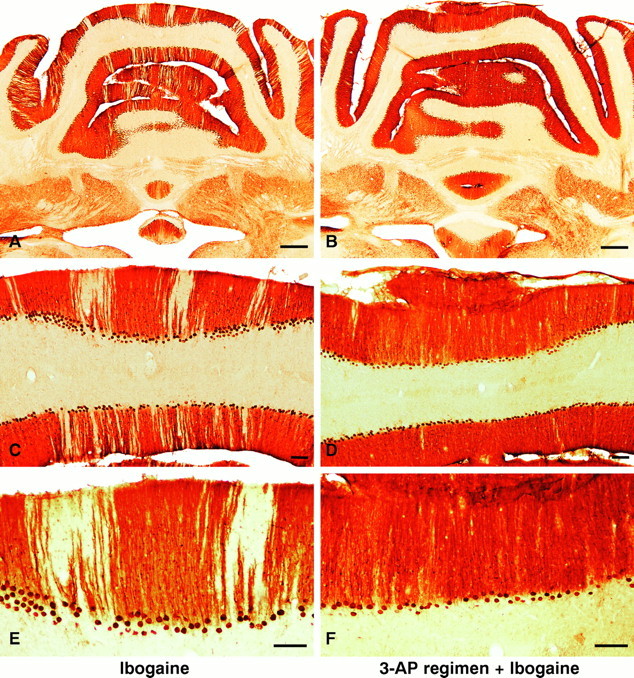
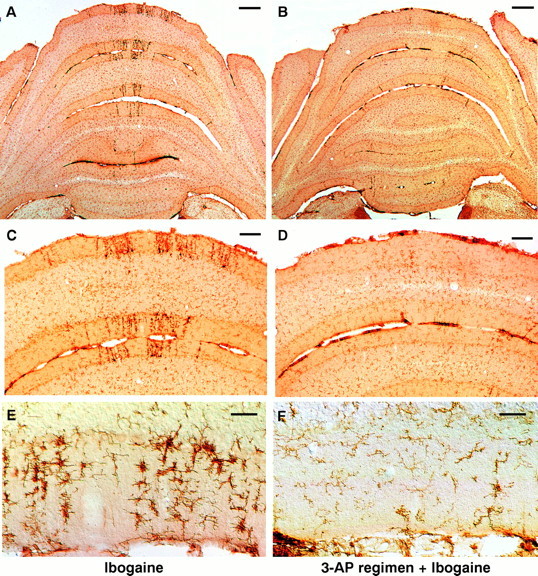
Ibogaine, an indole alkaloid that causes hallucinations, tremor, and ataxia, produces cerebellar neurotoxicity in rats, manifested by degeneration of Purkinje cells aligned in narrow parasagittal bands that are coextensive with activated glial cells. Harmaline, a closely related alkaloid that excites inferior olivary neurons, causes the same pattern of Purkinje cell degeneration, providing a clue to the mechanism of toxicity. We have proposed that ibogaine, like harmaline, excites neurons in the inferior olive, leading to sustained release of glutamate at climbing fiber synapses on Purkinje cells. The objective of this study was to test the hypothesis that increased climbing fiber activity induced by ibogaine mediates excitotoxic Purkinje cell degeneration. The inferior olive was pharmacologically ablated in rats by a neurotoxic drug regimen using 3-acetylpyridine, and cerebellar damage attributed to subsequent administration of ibogaine was analyzed using immunocytochemical markers for neurons and glial cells. The results show that ibogaine administered after inferior olive ablation produced little or no Purkinje cell degeneration or glial activation. That a lesion of the inferior olive almost completely prevents the neurotoxicity demonstrates that ibogaine is not directly toxic to Purkinje cells, but that the toxicity is indirect and dependent on integrity of the olivocerebellar projection. We postulate that ibogaine-induced activation of inferior olivary neurons leads to release of glutamate simultaneously at hundreds of climbing fiber terminals distributed widely over the surface of each Purkinje cell. The unique circuitry of the olivocerebellar projection provides this system with maximum synaptic security, a feature that confers on Purkinje cells a high degree of vulnerability to excitotoxic injury.
Figures
References
-
- Adams JH, Duchen LW. Greenfield’s neuropathology. Oxford UP; New York: 1992.
-
- Anderson WA, Flumerfelt BA. A light and electron microscopic study of the effects of 3-acetylpyridine intoxication on the inferior olivary complex and cerebellar cortex. J Comp Neurol. 1980;190:157–174. - PubMed
-
- Anderson WA, Flumerfelt BA. Sensitivity of rat inferior olivary neurons to 3-acetylpyridine. Dev Brain Res. 1984;12:285–291. - PubMed
-
- Azizi SA, Woodward DJ. Inferior olivary nuclear complex of the rat: morphology and comments on the principles of organization within the olivocerebellar system. J Comp Neurol. 1987;263:467–484. - PubMed
-
- Balaban CD. Central neurotoxic effects of intraperitoneally administered 3-acetylpyridine, harmaline and niacinamide in Sprague-Dawley and Long-Evans rats: a critical review of central 3-acetylpyridine neurotoxicity. Brain Res. 1985;356:21–42. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources