

Cell-adhesive responses to tenascin-C splice variants involve formation of fascin microspikes
- PMID: 9348542
- PMCID: PMC25670
- DOI: 10.1091/mbc.8.10.2055
Cell-adhesive responses to tenascin-C splice variants involve formation of fascin microspikes
Abstract
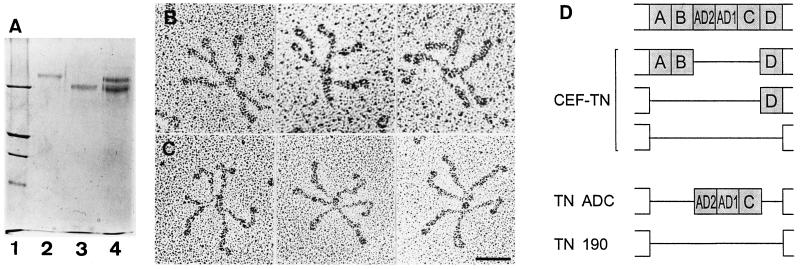
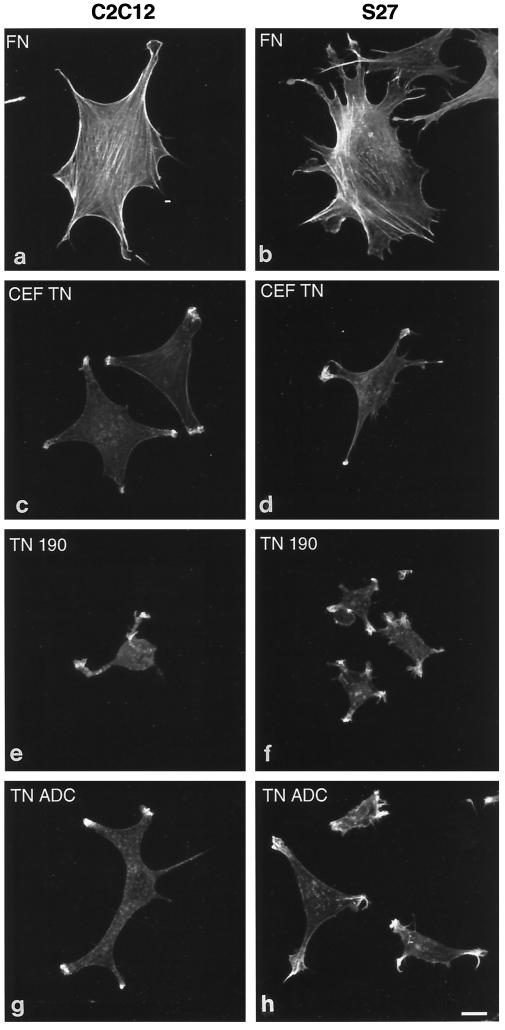
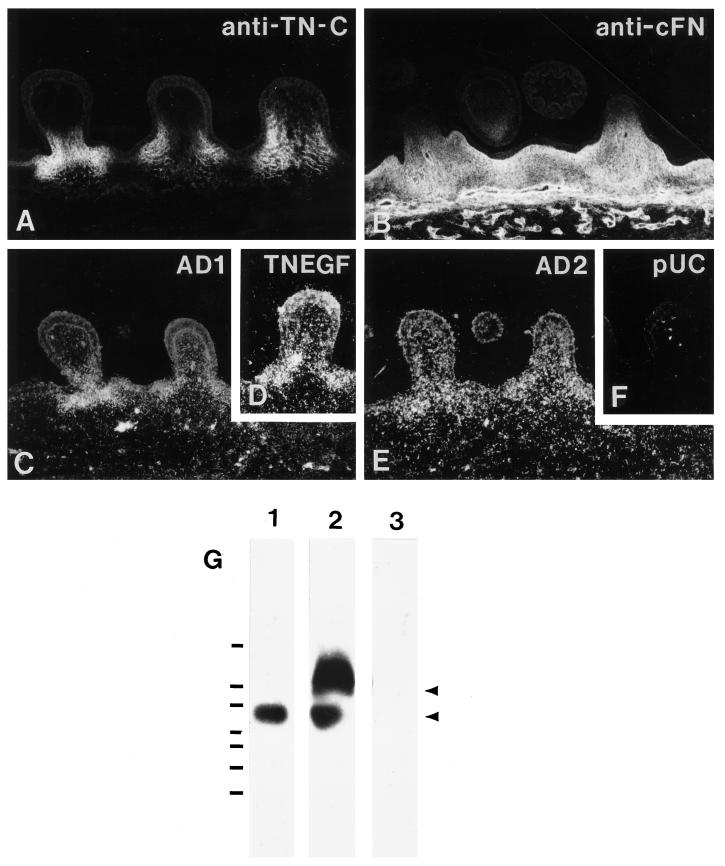
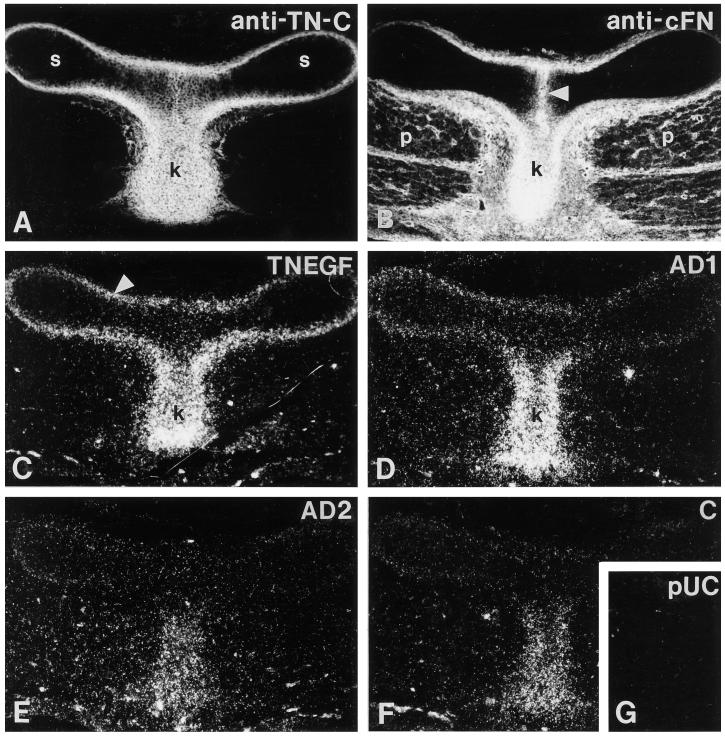
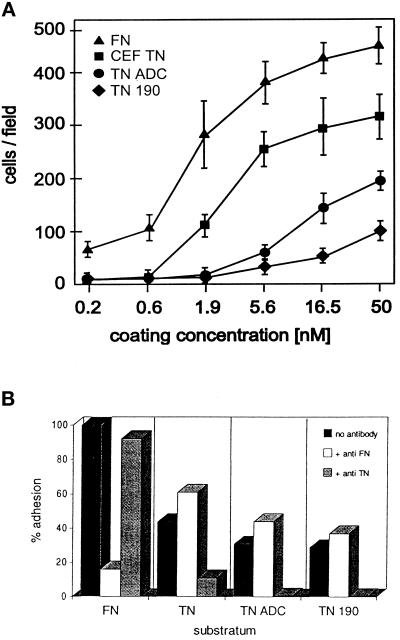
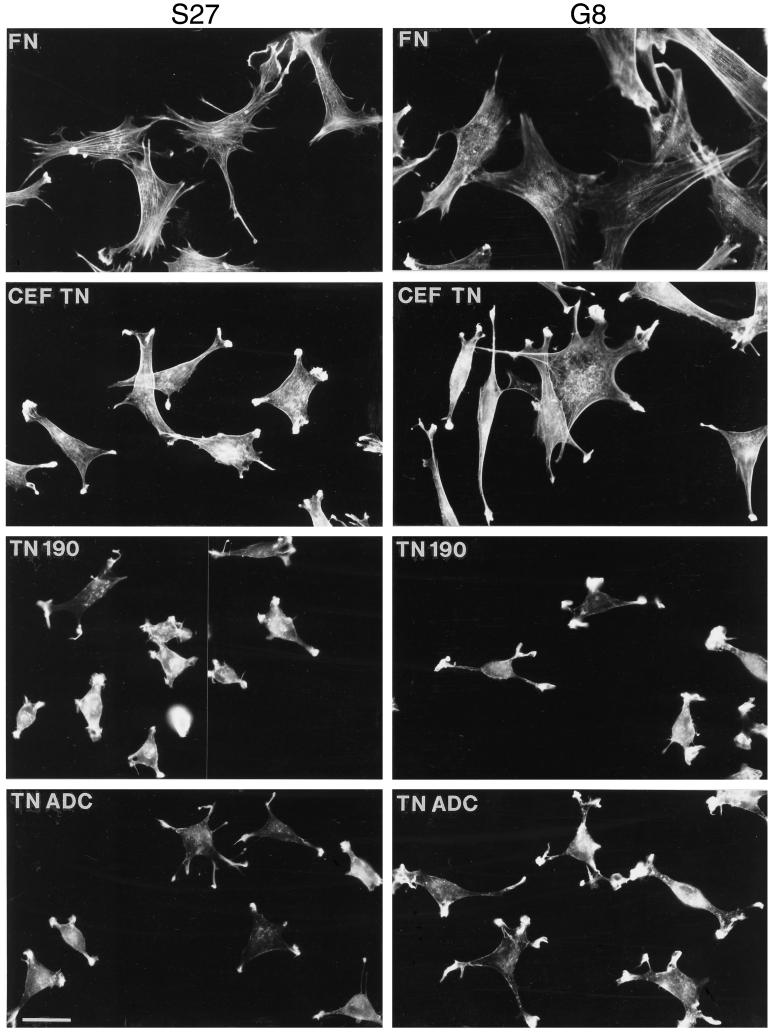
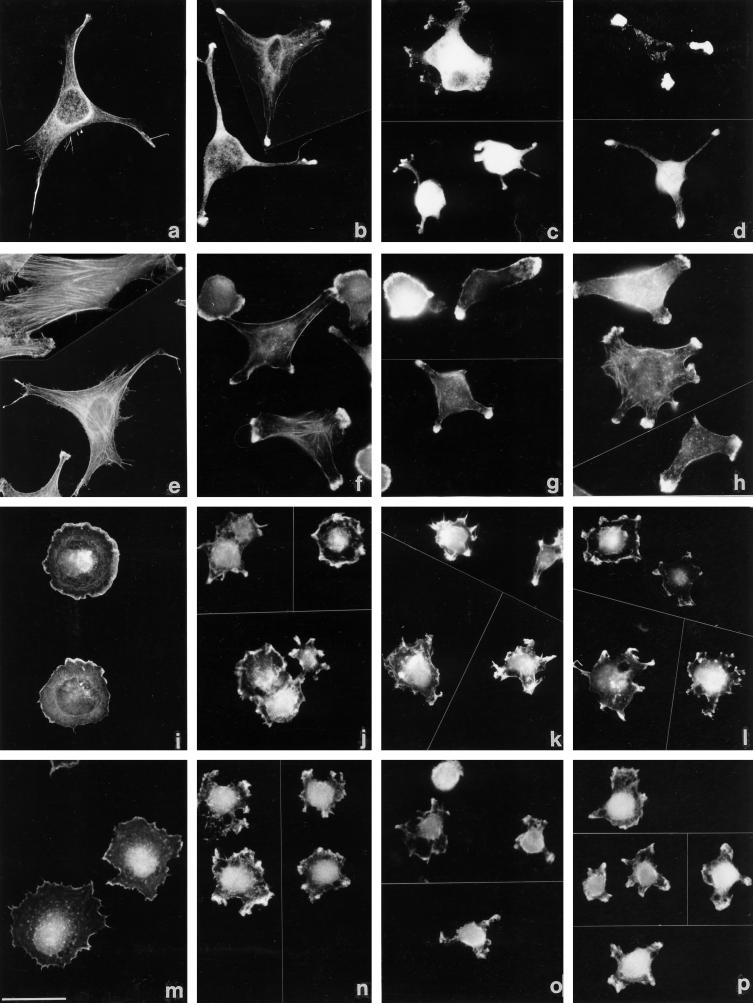
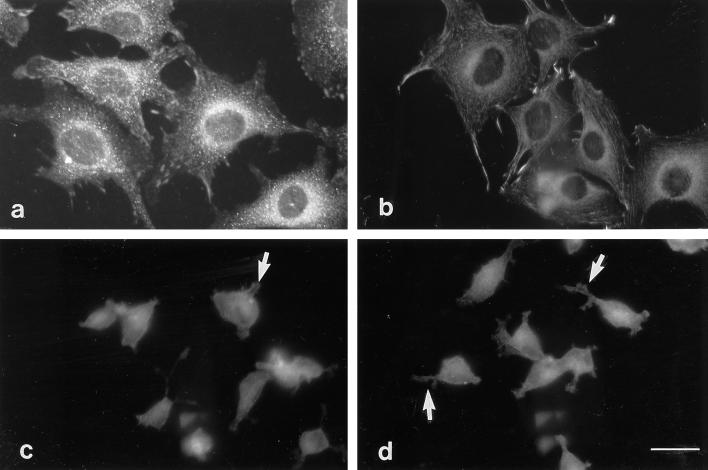
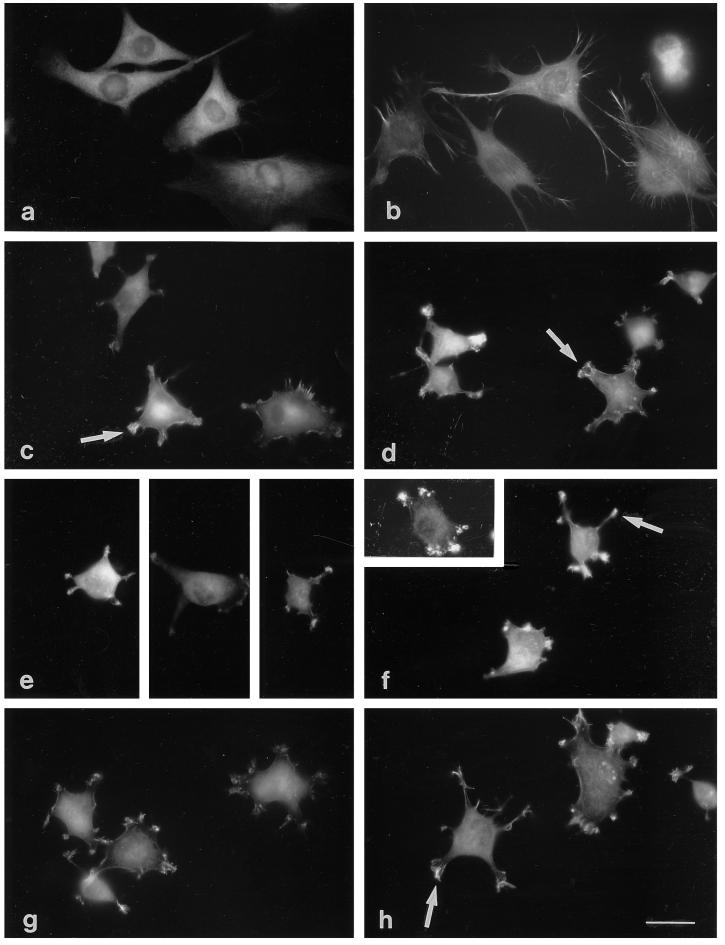
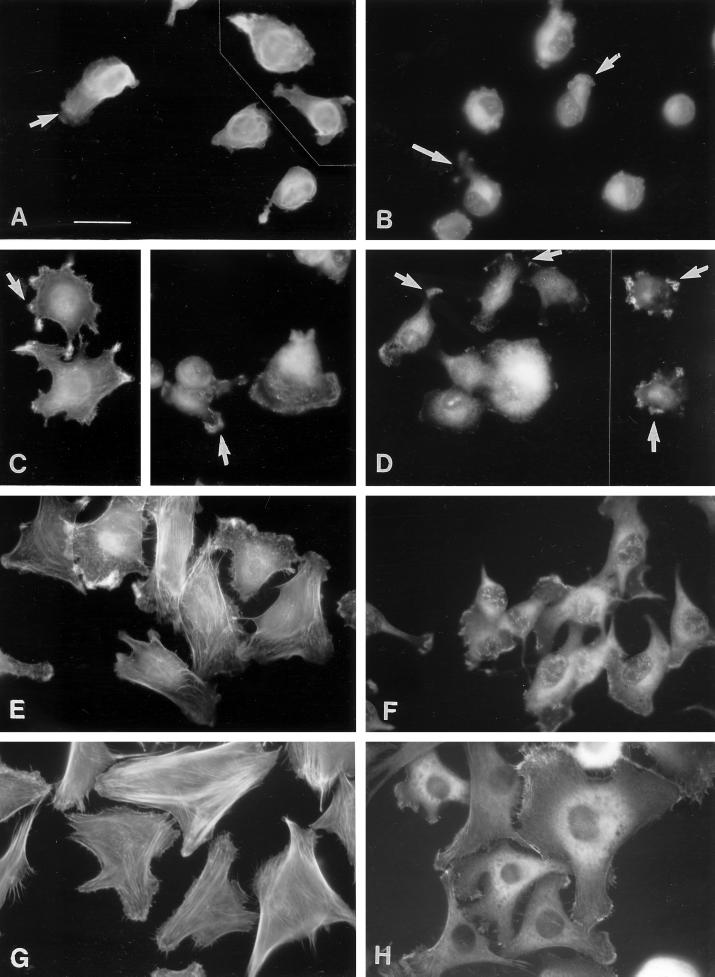
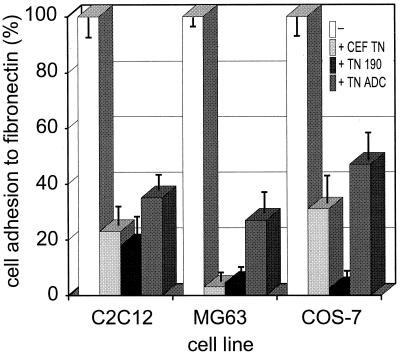
Tenascin-C is an adhesion-modulating matrix glycoprotein that has multiple effects on cell behavior. Tenascin-C transcripts are expressed in motile cells and at sites of tissue modeling during development, and alternative splicing generates variants that encode different numbers of fibronectin type III repeats. We have examined the in vivo expression and cell adhesive properties of two full-length recombinant tenascin-C proteins: TN-190, which contains the eight constant fibronectin type III repeats, and TN-ADC, which contains the additional AD2, AD1, and C repeats. In situ hybridization with probes specific for the AD2, AD1, and C repeats shows that these splice variants are expressed at sites of active tissue modeling and fibronectin expression in the developing avian feather bud and sternum. Transcripts incorporating the AD2, AD1, and C repeats are present in embryonic day 10 wing bud but not in embryonic day 10 lung. By using a panel of nine cell lines in attachment assays, we have found that C2C12, G8, and S27 myoblastic cells undergo concentration-dependent adhesion to both variants, organize actin microspikes that contain the actin-bundling protein fascin, and do not assemble focal contacts. On a molar basis, TN-ADC is more active than TN-190 in promoting cell attachment and irregular cell spreading. The addition of either TN-190 or TN-ADC in solution to C2C12, COS-7, or MG-63 cells adherent on fibronectin decreases cell attachment and results in decreased organization of actin microfilament bundles, with formation of cortical membrane ruffles and retention of residual points of substratum contact that contain filamentous actin and fascin. These data establish a biochemical similarity in the processes of cell adhesion to tenascin-C and thrombospondin-1, also an "antiadhesive" matrix component, and also demonstrate that both the adhesive and adhesion-modulating properties of tenascin-C involve similar biochemical events in the cortical cytoskeleton. In addition to these generic properties, TN-ADC is less active in adhesion modulation than TN-190. The coordinated expression of different tenascin-C transcripts during development may, therefore, provide appropriate microenvironments for regulated changes in cell shape, adhesion, and movement.
Figures
References
-
- Adams JC. Formation of stable microspikes containing actin and the 55-kDa actin bundling protein, fascin, is a consequence of cell adhesion to thrombospondin-1: implications for the anti-adhesive activities of thrombospondin-1. J Cell Sci. 1995;108:1977–1990. - PubMed
-
- Adams JC, Lawler J. Diverse mechanisms for cell attachment to platelet thrombospondin. J Cell Sci. 1993;104:1061–1071. - PubMed
-
- Adams JC, Tucker RP, Lawler J. The Thrombospondin Gene Family. Austin, TX: R.G. Landes; 1995. and Berlin: Springer-Verlag.
-
- Asch AS, Tepler J, Silbiger S, Nachman RL. Cellular attachment to thrombospondin: cooperative interactions between receptor systems. J Biol Chem. 1991;266:1740–1745. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
