

The involvement of the intermediate chain of cytoplasmic dynein in binding the motor complex to membranous organelles of Xenopus oocytes
- PMID: 9348543
- PMCID: PMC25673
- DOI: 10.1091/mbc.8.10.2077
The involvement of the intermediate chain of cytoplasmic dynein in binding the motor complex to membranous organelles of Xenopus oocytes
Abstract
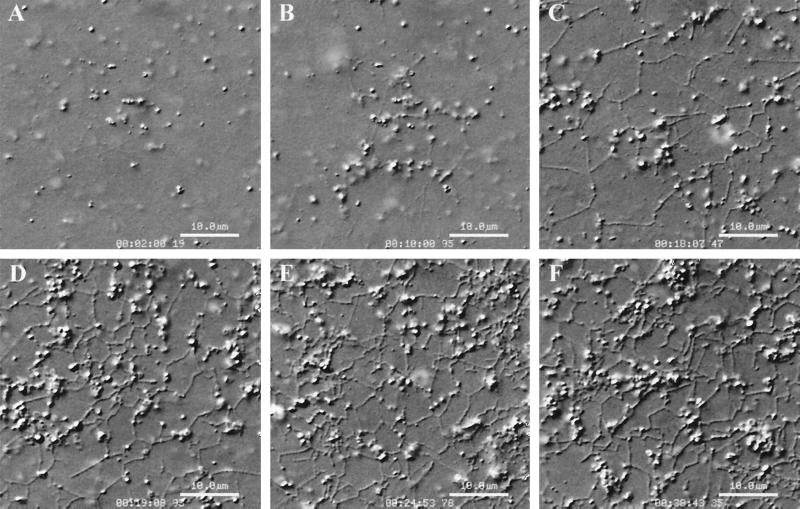
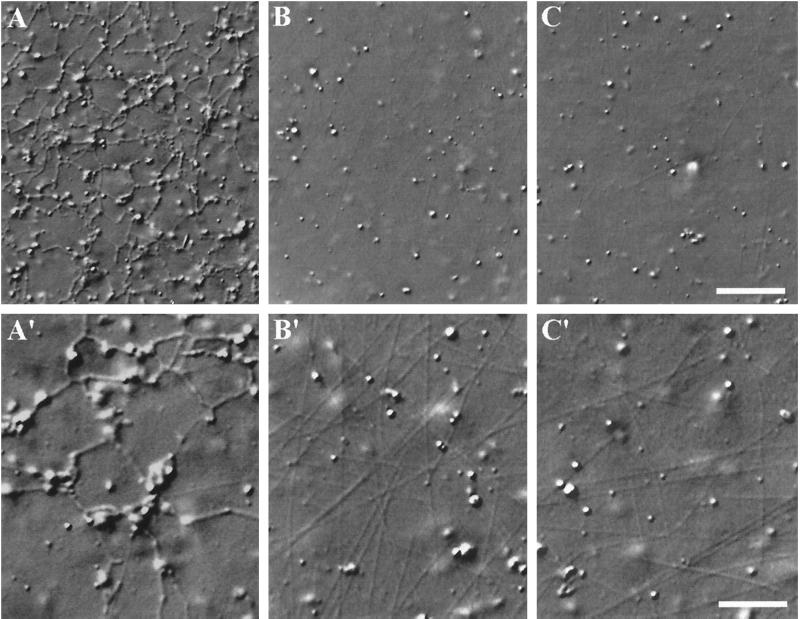
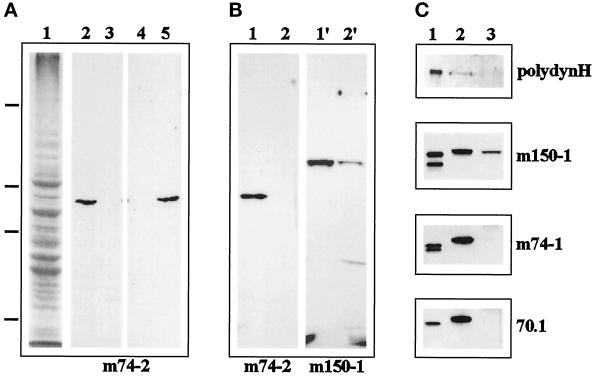
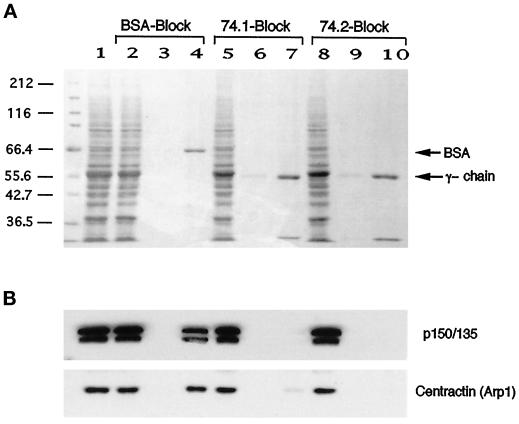
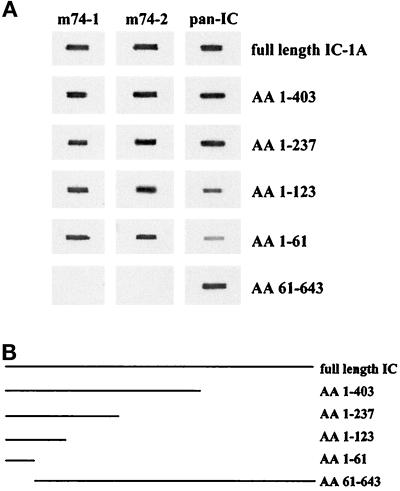
Cytoplasmic dynein is one of the major motor proteins involved in intracellular transport. It is a protein complex consisting of four subunit classes: heavy chains, intermediate chains (ICs), light intermediate chains, and light chains. In a previous study, we had generated new monoclonal antibodies to the ICs and mapped the ICs to the base of the motor. Because the ICs have been implicated in targeting the motor to cargo, we tested whether these new antibodies to the intermediate chain could block the function of cytoplasmic dynein. When cytoplasmic extracts of Xenopus oocytes were incubated with either one of the monoclonal antibodies (m74-1, m74-2), neither organelle movement nor network formation was observed. Network formation and membrane transport was blocked at an antibody concentration as low as 15 micrograms/ml. In contrast to these observations, no effect was observed on organelle movement and tubular network formation in the presence of a control antibody at concentrations as high as 0.5 mg/ml. After incubating cytoplasmic extracts or isolated membranes with the monoclonal antibodies m74-1 and m74-2, the dynein IC polypeptide was no longer detectable in the membrane fraction by SDS-PAGE immunoblot, indicating a loss of cytoplasmic dynein from the membrane. We used a panel of dynein IC truncation mutants and mapped the epitopes of both antibodies to the N-terminal coiled-coil domain, in close proximity to the p150Glued binding domain. In an IC affinity column binding assay, both antibodies inhibited the IC-p150Glued interaction. Thus these findings demonstrate that direct IC-p150Glued interaction is required for the proper attachment of cytoplasmic dynein to membranes.
Figures

Similar articles
-
Apoptotic cleavage of cytoplasmic dynein intermediate chain and p150(Glued) stops dynein-dependent membrane motility.J Cell Biol. 2001 Jun 25;153(7):1415-26. doi: 10.1083/jcb.153.7.1415. J Cell Biol. 2001. PMID: 11425872 Free PMC article.
-
Cytoplasmic dynein binds dynactin through a direct interaction between the intermediate chains and p150Glued.J Cell Biol. 1995 Dec;131(6 Pt 1):1507-16. doi: 10.1083/jcb.131.6.1507. J Cell Biol. 1995. PMID: 8522607 Free PMC article.
-
Affinity chromatography demonstrates a direct binding between cytoplasmic dynein and the dynactin complex.J Biol Chem. 1995 Dec 1;270(48):28806-11. doi: 10.1074/jbc.270.48.28806. J Biol Chem. 1995. PMID: 7499404
-
Regulation of dynein-dynactin-driven vesicular transport.Traffic. 2017 Jun;18(6):336-347. doi: 10.1111/tra.12475. Epub 2017 Mar 28. Traffic. 2017. PMID: 28248450 Review.
-
Targeting of motor proteins.Science. 1996 Mar 15;271(5255):1539-44. doi: 10.1126/science.271.5255.1539. Science. 1996. PMID: 8599110 Review.
Cited by
-
The axonal transport of mitochondria.J Cell Sci. 2012 May 1;125(Pt 9):2095-104. doi: 10.1242/jcs.053850. Epub 2012 May 22. J Cell Sci. 2012. PMID: 22619228 Free PMC article. Review.
-
A molecular genetic analysis of the interaction between the cytoplasmic dynein intermediate chain and the glued (dynactin) complex.Mol Biol Cell. 2000 Nov;11(11):3791-803. doi: 10.1091/mbc.11.11.3791. Mol Biol Cell. 2000. PMID: 11071907 Free PMC article.
-
The kinesin KIF1C and microtubule plus ends regulate podosome dynamics in macrophages.Mol Biol Cell. 2006 Jun;17(6):2811-23. doi: 10.1091/mbc.e05-11-1010. Epub 2006 Mar 22. Mol Biol Cell. 2006. PMID: 16554367 Free PMC article.
-
Optimizing Efficiency and Motility of a Polyvalent Molecular Motor.Micromachines (Basel). 2022 Jun 9;13(6):914. doi: 10.3390/mi13060914. Micromachines (Basel). 2022. PMID: 35744528 Free PMC article.
-
Bidirectional translocation of neurofilaments along microtubules mediated in part by dynein/dynactin.Mol Biol Cell. 2000 Oct;11(10):3495-508. doi: 10.1091/mbc.11.10.3495. Mol Biol Cell. 2000. PMID: 11029051 Free PMC article.
References
-
- Adams RJ, Pollard TD. Binding of myosin I to membrane lipids. Nature. 1989;340:565–568. - PubMed
-
- Allan V. Assay of membrane motility in interphase and metaphase Xenopus extracts. Methods Cell Biol. 1993;39:203–226. - PubMed
-
- Allan V, Vale R. Movement of membrane tubules along microtubules in vitro: evidence for specialized sites of motor attachment. J Cell Sci. 1994;107:1885–1897. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
