

Inhibition of ubiquitin/proteasome-dependent protein degradation by the Gly-Ala repeat domain of the Epstein-Barr virus nuclear antigen 1
- PMID: 9356498
- PMCID: PMC25057
- DOI: 10.1073/pnas.94.23.12616
Inhibition of ubiquitin/proteasome-dependent protein degradation by the Gly-Ala repeat domain of the Epstein-Barr virus nuclear antigen 1
Abstract
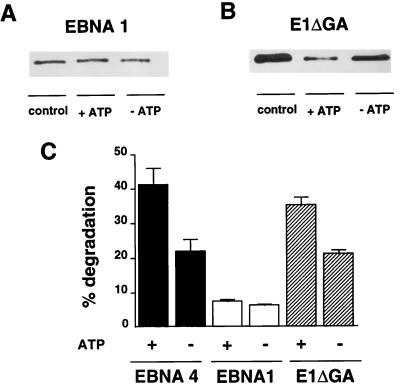
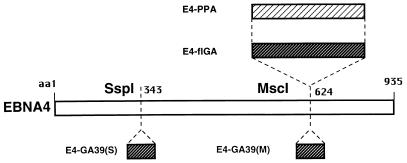
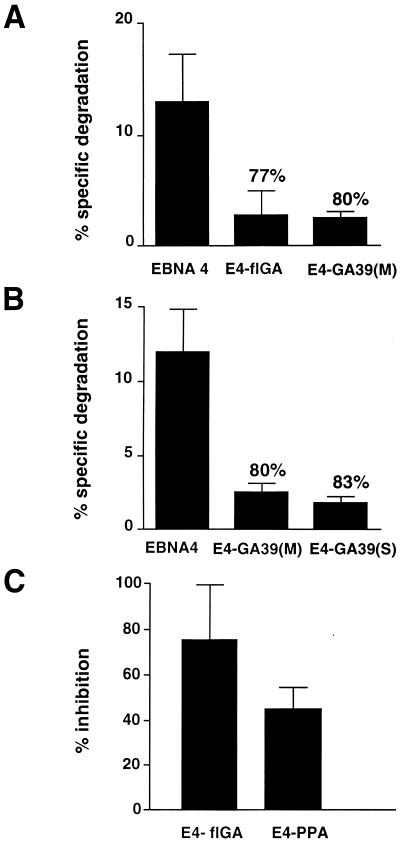
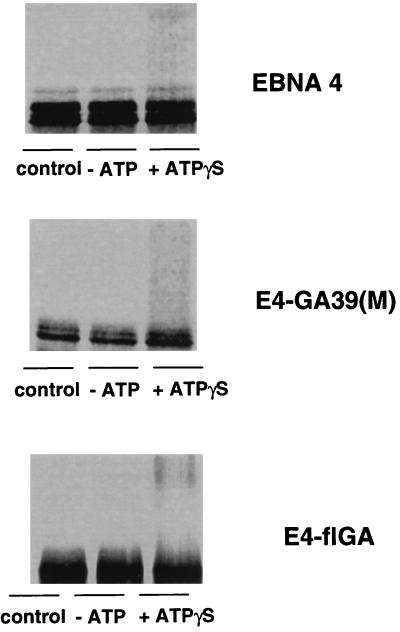
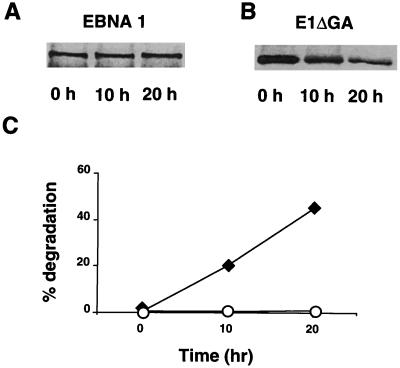
The Epstein-Barr virus (EBV) encoded nuclear antigen (EBNA) 1 is expressed in latently infected B lymphocytes that persist for life in healthy virus carriers and is the only viral protein regularly detected in all EBV associated malignancies. The Gly-Ala repeat domain of EBNA1 was shown to inhibit in cis the presentation of major histocompatibility complex (MHC) class I restricted cytotoxic T cell epitopes from EBNA4. It appears that the majority of antigens presented via the MHC I pathway are subject to ATP-dependent ubiquitination and degradation by the proteasome. We have investigated the influence of the repeat on this process by comparing the degradation of EBNA1, EBNA4, and Gly-Ala containing EBNA4 chimeras in a cell-free system. EBNA4 was efficiently degraded in an ATP/ubiquitin/proteasome-dependent fashion whereas EBNA1 was resistant to degradation. Processing of EBNA1 was restored by deletion of the Gly-Ala domain whereas insertion of Gly-Ala repeats of various lengths and in different positions prevented the degradation of EBNA4 without appreciable effect on ubiquitination. Inhibition was also achieved by insertion of a Pro-Ala coding sequence. The results suggest that the repeat may affect MHC I restricted responses by inhibiting antigen processing via the ubiquitin/proteasome pathway. The presence of regularly interspersed Ala residues appears to be important for the effect.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
