

Degeneration of neurons, synapses, and neuropil and glial activation in a murine Atm knockout model of ataxia-telangiectasia
- PMID: 9356511
- PMCID: PMC25086
- DOI: 10.1073/pnas.94.23.12688
Degeneration of neurons, synapses, and neuropil and glial activation in a murine Atm knockout model of ataxia-telangiectasia
Abstract
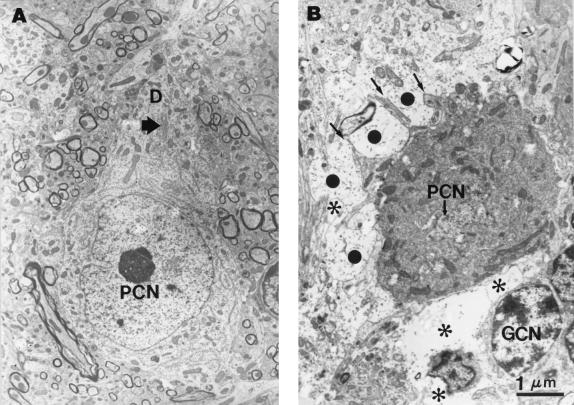
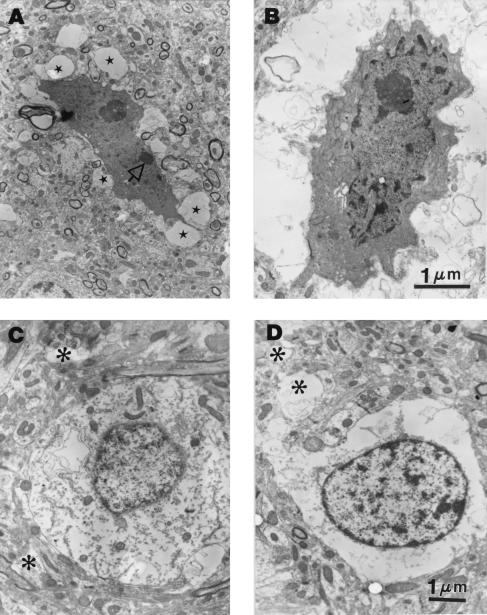
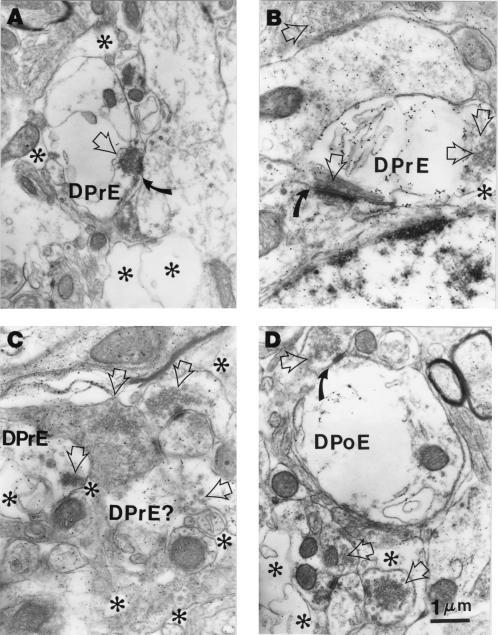
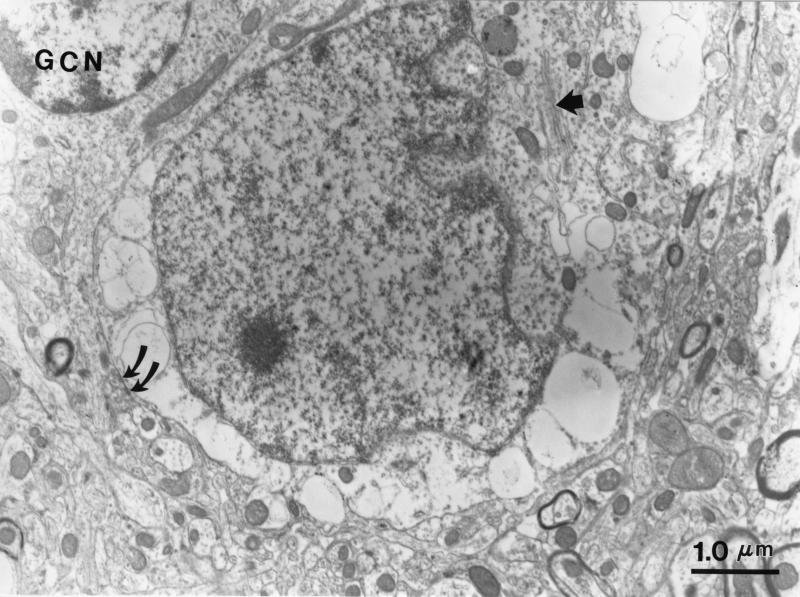
Neural degeneration is one of the clinical manifestations of ataxia-telangiectasia, a disorder caused by mutations in the Atm protein kinase gene. However, neural degeneration was not detected with general purpose light microscopic methods in previous studies using several different lines of mice with disrupted Atm genes. Here, we show electron microscopic evidence of degeneration of several different types of neurons in the cerebellar cortex of 2-month-old Atm knockout mice, which is accompanied by glial activation, deterioration of neuropil structure, and both pre- and postsynaptic degeneration. These findings are similar to those in patients with ataxia-telangiectasia, indicating that Atm knockout mice are a useful model to elucidate the mechanisms underlying neurodegeneration in this condition and to develop and test strategies to palliate and prevent the disease.
Figures
Similar articles
-
Mutations in String/CDC25 inhibit cell cycle re-entry and neurodegeneration in a Drosophila model of Ataxia telangiectasia.Genes Dev. 2008 May 1;22(9):1205-20. doi: 10.1101/gad.1639608. Epub 2008 Apr 11. Genes Dev. 2008. PMID: 18408079 Free PMC article.
-
The innate immune response transcription factor relish is necessary for neurodegeneration in a Drosophila model of ataxia-telangiectasia.Genetics. 2013 May;194(1):133-42. doi: 10.1534/genetics.113.150854. Epub 2013 Mar 15. Genetics. 2013. PMID: 23502677 Free PMC article.
-
Activation of AMP-activated protein kinase in cerebella of Atm-/- mice is attributable to accumulation of reactive oxygen species.Biochem Biophys Res Commun. 2012 Feb 10;418(2):267-72. doi: 10.1016/j.bbrc.2012.01.008. Epub 2012 Jan 10. Biochem Biophys Res Commun. 2012. PMID: 22260947 Free PMC article.
-
ATM: the product of the gene mutated in ataxia-telangiectasia.Int J Biochem Cell Biol. 1999 Jul;31(7):735-40. doi: 10.1016/s1357-2725(99)00028-x. Int J Biochem Cell Biol. 1999. PMID: 10467728 Review.
-
ATM: the protein encoded by the gene mutated in the radiosensitive syndrome ataxia-telangiectasia.Int J Radiat Biol. 1999 Oct;75(10):1201-14. doi: 10.1080/095530099139359. Int J Radiat Biol. 1999. PMID: 10549596 Review.
Cited by
-
SMRT compounds abrogate cellular phenotypes of ataxia telangiectasia in neural derivatives of patient-specific hiPSCs.Nat Commun. 2013;4:1824. doi: 10.1038/ncomms2824. Nat Commun. 2013. PMID: 23652012
-
SEQUIN Multiscale Imaging of Mammalian Central Synapses Reveals Loss of Synaptic Connectivity Resulting from Diffuse Traumatic Brain Injury.Neuron. 2020 Jul 22;107(2):257-273.e5. doi: 10.1016/j.neuron.2020.04.012. Epub 2020 May 8. Neuron. 2020. PMID: 32392471 Free PMC article.
-
DNA repair abnormalities leading to ataxia: shared neurological phenotypes and risk factors.Neurogenetics. 2014 Oct;15(4):217-28. doi: 10.1007/s10048-014-0415-z. Epub 2014 Jul 20. Neurogenetics. 2014. PMID: 25038946 Review.
-
Involvement of p53 and p21 in cellular defects and tumorigenesis in Atm-/- mice.Mol Cell Biol. 1998 Jul;18(7):4385-90. doi: 10.1128/MCB.18.7.4385. Mol Cell Biol. 1998. PMID: 9632822 Free PMC article.
-
LINE-1 activation in the cerebellum drives ataxia.Neuron. 2022 Oct 19;110(20):3278-3287.e8. doi: 10.1016/j.neuron.2022.08.011. Epub 2022 Sep 6. Neuron. 2022. PMID: 36070749 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
