

Integrin alpha 6A beta 1 induces CD81-dependent cell motility without engaging the extracellular matrix migration substrate
- PMID: 9362067
- PMCID: PMC25706
- DOI: 10.1091/mbc.8.11.2253
Integrin alpha 6A beta 1 induces CD81-dependent cell motility without engaging the extracellular matrix migration substrate
Abstract
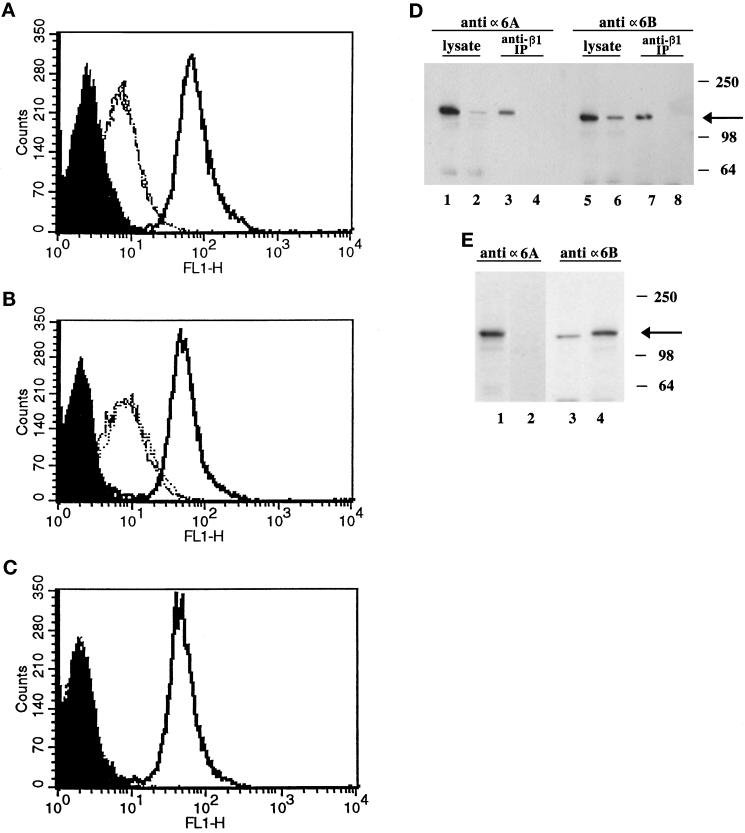
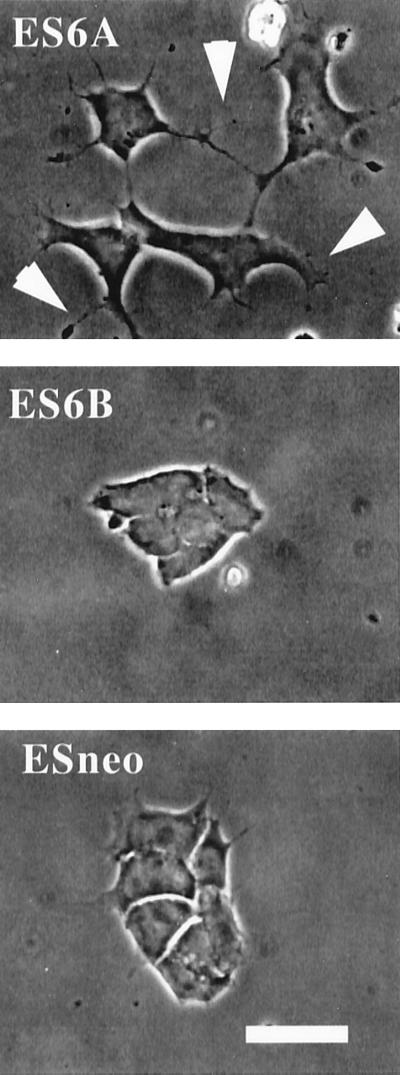
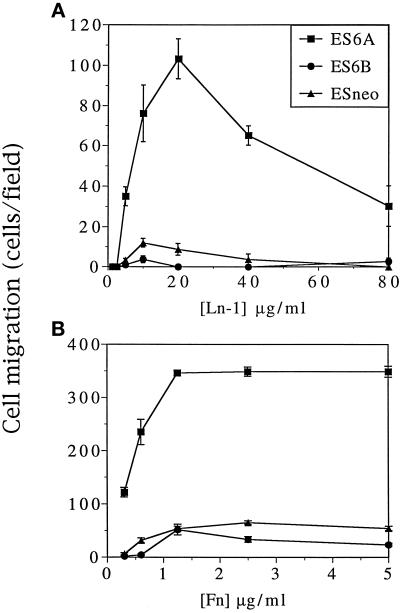
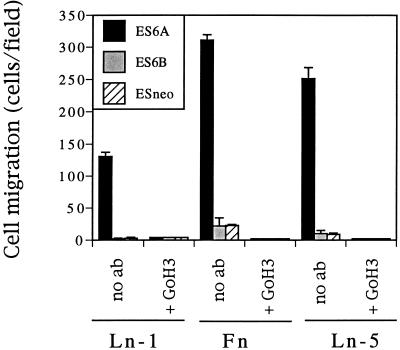
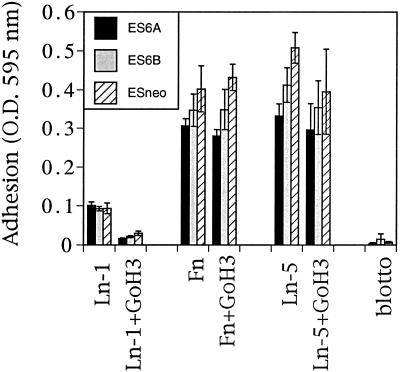
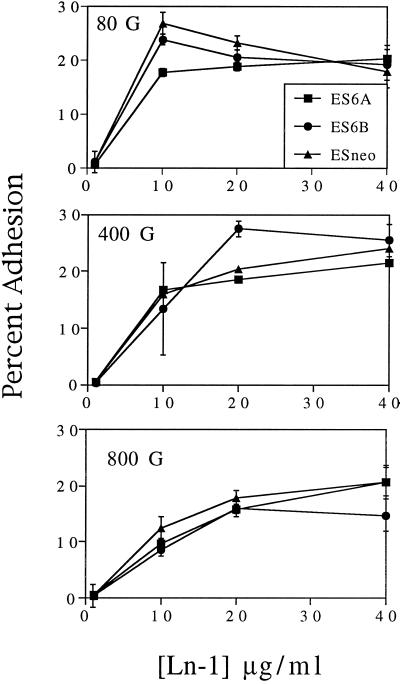
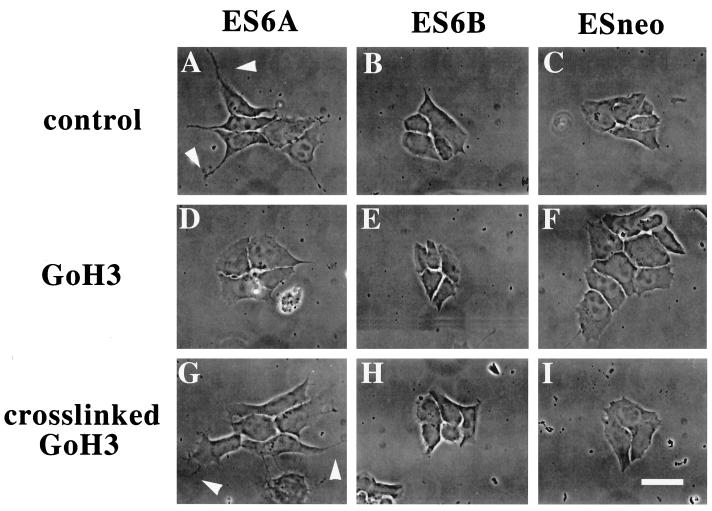
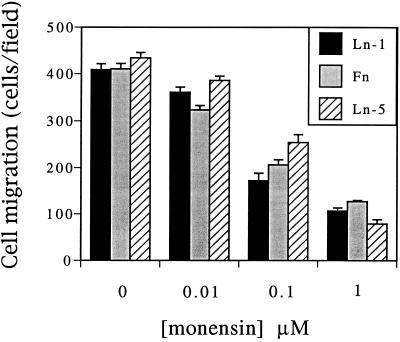
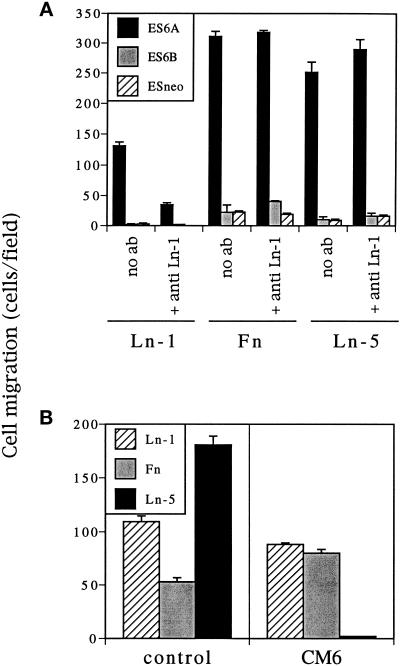
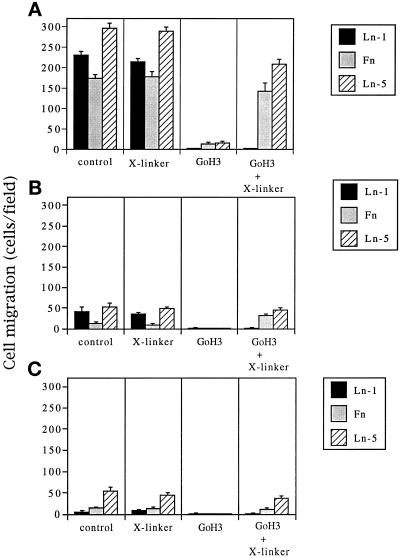
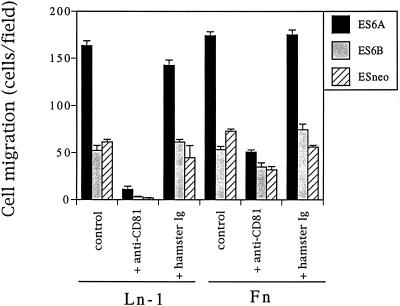
It is well established that integrins and extracellular matrix (ECM) play key roles in cell migration, but the underlying mechanisms are poorly defined. We describe a novel mechanism whereby the integrin alpha 6 beta 1, a laminin receptor, can affect cell motility and induce migration onto ECM substrates with which it is not engaged. By using DNA-mediated gene transfer, we expressed the human integrin subunit alpha 6A in murine embryonic stem (ES) cells. ES cells expressing alpha 6A (ES6A) at the surface dimerized with endogenous beta 1, extended numerous filopodia and lamellipodia, and were intensely migratory in haptotactic assays on laminin (LN)-1. Transfected alpha 6A was responsible for these effects, because cells transfected with control vector or alpha 6B, a cytoplasmic domain alpha 6 isoform, displayed compact morphology and no migration, like wild-type ES cells. The ES6A migratory phenotype persisted on fibronectin (Fn) and Ln-5. Adhesion inhibition assays indicated that alpha 6 beta 1 did not contribute detectably to adhesion to these substrates in ES cells. However, anti-alpha 6 antibodies completely blocked migration of ES6A cells on Fn or Ln-5. Control experiments with monensin and anti-ECM antibodies indicated that this inhibition could not be explained by deposition of an alpha 6 beta 1 ligand (e.g., Ln-1) by ES cells. Cross-linking with secondary antibody overcame the inhibitory effect of anti-alpha 6 antibodies, restoring migration or filopodia extension on Fn and Ln-5. Thus, to induce migration in ES cells, alpha 6A beta 1 did not have to engage with an ECM ligand but likely participated in molecular interactions sensitive to anti-alpha 6 beta 1 antibody and mimicked by cross-linking. Antibodies to the tetraspanin CD81 inhibited alpha 6A beta 1-induced migration but had no effect on ES cell adhesion. It is known that CD81 is physically associated with alpha 6 beta 1, therefore our results suggest a mechanism by which interactions between alpha 6A beta 1 and CD81 may up-regulate cell motility, affecting migration mediated by other integrins.
Figures

References
-
- Abe, T., Ohno, M., Sato, T., Murakami, M., Kajiki, M., and Kodaira, R. (1991). “Differentiation induction” culture of human leukemic myeloid cells stimulates high production of macrophage differentiation inducing factor. [Review]. Cytotechnology 5(suppl 2), S75–S93. - PubMed
-
- Boismenu R, Rhein M, Fischer WH, Havran WL. A role for CD81 in early T cell development. Science. 1996;271:198–200. - PubMed
-
- Chan BM, Kassner PD, Schiro JA, Byers HR, Kupper TS, Hemler ME. Distinct cellular functions mediated by different VLA integrin alpha subunit cytoplasmic domains. Cell. 1992;68:1051–1060. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
