

End4p/Sla2p interacts with actin-associated proteins for endocytosis in Saccharomyces cerevisiae
- PMID: 9362070
- PMCID: PMC25709
- DOI: 10.1091/mbc.8.11.2291
End4p/Sla2p interacts with actin-associated proteins for endocytosis in Saccharomyces cerevisiae
Abstract
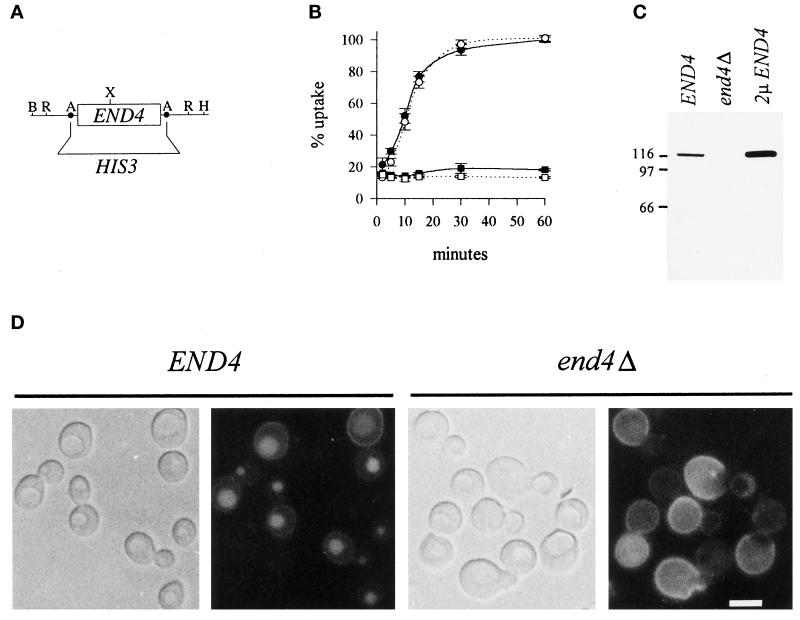
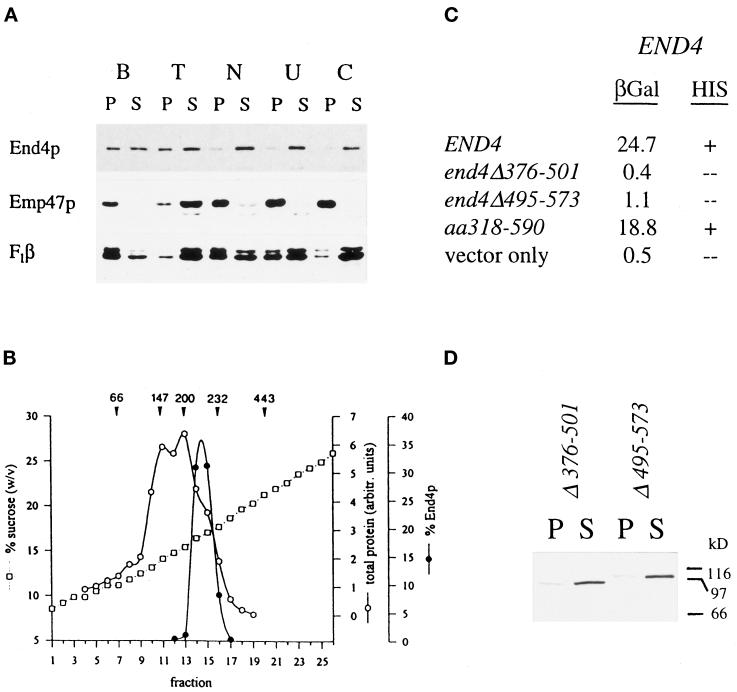
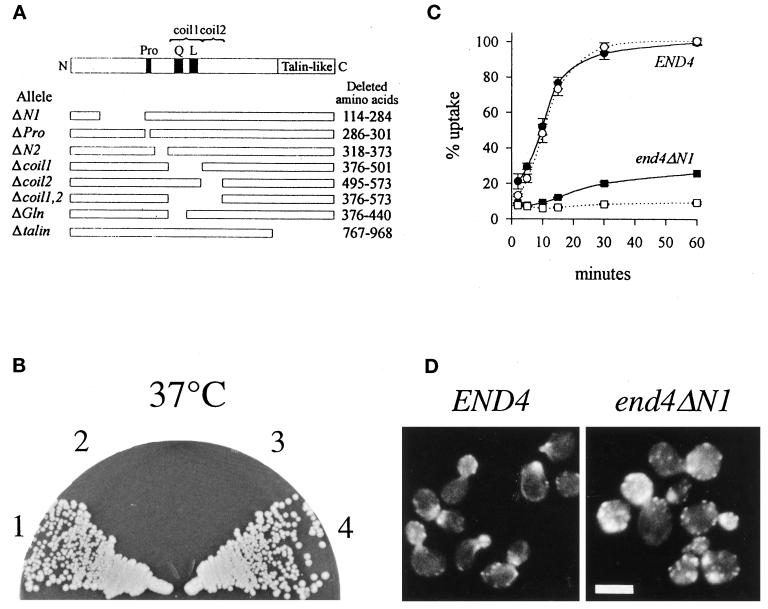
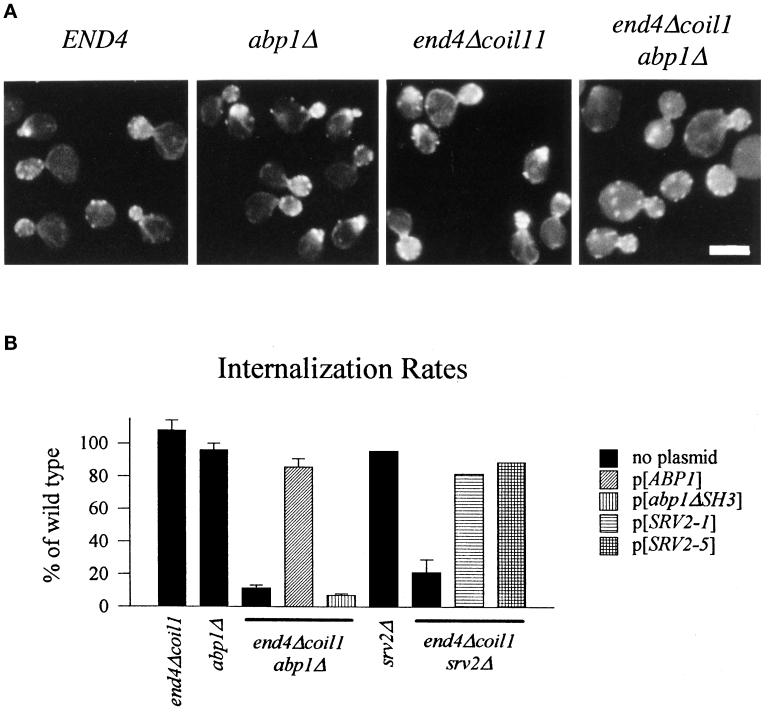
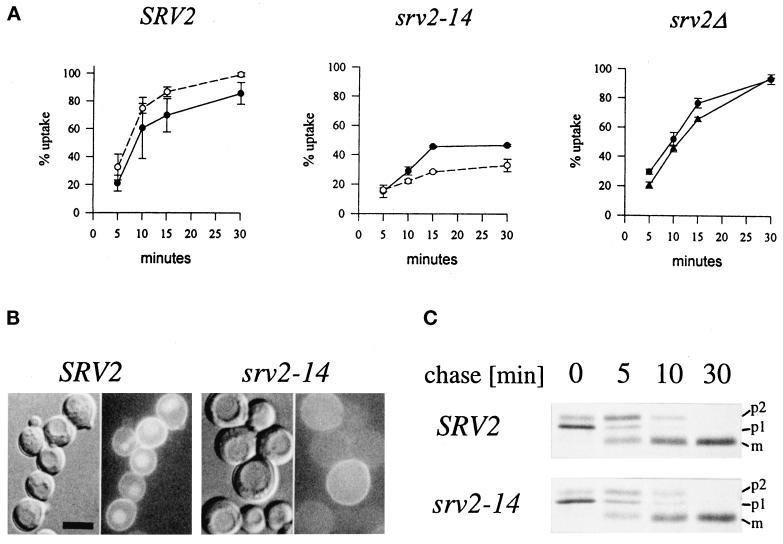
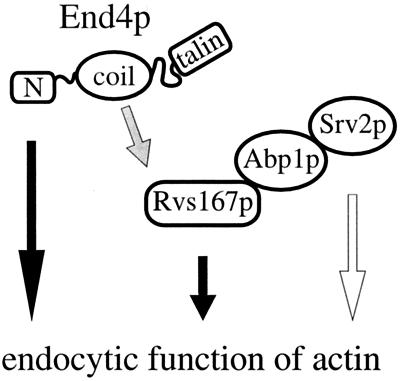
end4-1 was isolated as a temperature-sensitive endocytosis mutant. We cloned and sequenced END4 and found that it is identical to SLA2/MOP2. This gene is required for growth at high temperature, viability in the absence of Abp1p, polarization of the cortical actin cytoskeleton, and endocytosis. We used a mutational analysis of END4 to correlate in vivo functions with regions of End4p and we found that two regions of End4p participate in endocytosis but that the talin-like domain of End4p is dispensable. The N-terminal domain of End4p is required for growth at high temperature, endocytosis, and actin organization. A central coiled-coil domain of End4p is necessary for formation of a soluble sedimentable complex. Furthermore, this domain has an endocytic function that is redundant with the function(s) of ABP1 and SRV2. The endocytic function of Abp1p depends on its SH3 domain. In addition we have isolated a recessive negative allele of SRV2 that is defective for endocytosis. Combined biochemical, functional, and genetic analysis lead us to propose that End4p may mediate endocytosis through interaction with other actin-associated proteins, perhaps Rvs167p, a protein essential for endocytosis.
Figures
References
-
- Burridge K, Fath K, Kelly T, Nuckolls G, Turner C. Focal adhesions: transmembrane junctions between the extracellular matrix and the cytoskeleton. Annu Rev Cell Biol. 1988;4:487–525. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
