

Precocious mammary gland development in P-cadherin-deficient mice
- PMID: 9362520
- PMCID: PMC2139972
- DOI: 10.1083/jcb.139.4.1025
Precocious mammary gland development in P-cadherin-deficient mice
Abstract
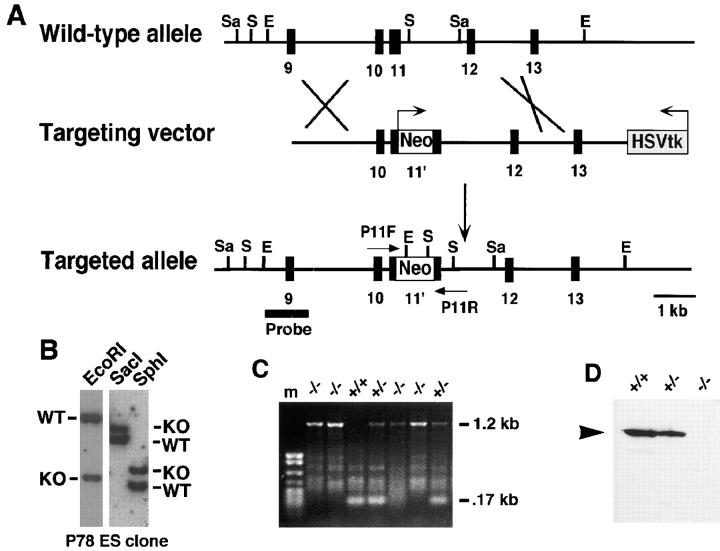
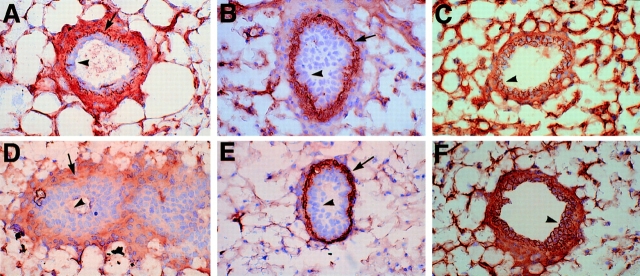
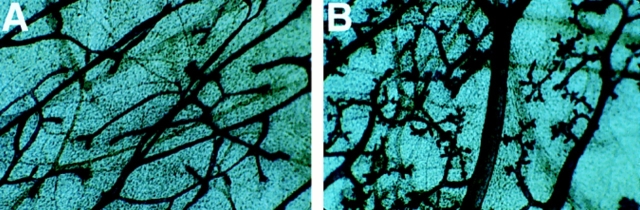
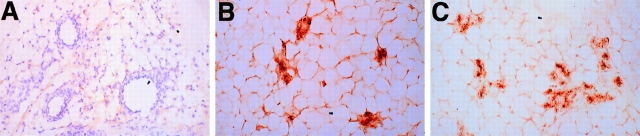
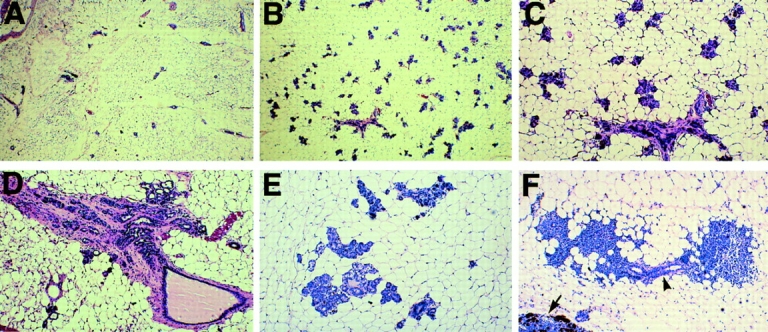
To investigate the functions of P-cadherin in vivo, we have mutated the gene encoding this cell adhesion receptor in mice. In contrast to E- and N-cadherin- deficient mice, mice homozygous for the P-cadherin mutation are viable. Although P-cadherin is expressed at high levels in the placenta, P-cadherin-null females are fertile. P-cadherin expression is localized to the myoepithelial cells surrounding the lumenal epithelial cells of the mammary gland. The role of the myoepithelium as a contractile tissue necessary for milk secretion is clear, but its function in the nonpregnant animal is unknown. The ability of the P-cadherin mutant female to nurse and maintain her litter indicates that the contractile function of the myoepithelium is not dependent on the cell adhesion molecule P-cadherin. The virgin P-cadherin-null females display precocious differentiation of the mammary gland. The alveolar-like buds in virgins resemble the glands of an early pregnant animal morphologically and biochemically (i.e., milk protein synthesis). The P-cadherin mutant mice develop hyperplasia and dysplasia of the mammary epithelium with age. In addition, abnormal lymphocyte infiltration was observed in the mammary glands of the mutant animals. These results indicate that P-cadherin-mediated adhesion and/or signals derived from cell-cell interactions are important determinants in negative growth control in the mammary gland. Furthermore, the loss of P-cadherin from the myoepithelium has uncovered a novel function for this tissue in maintaining the undifferentiated state of the underlying secretory epithelium.
Figures
References
-
- Bchini O, Andres AC, Schubaur B, Mehtali M, LeMeur M, Lathe R, Gerlinger P. Precocious mammary gland development and milk protein synthesis in transgenic mice ubiquitously expressing human growth hormone. Endocrinology. 1991;128:539–546. - PubMed
-
- Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature. 1996;382:638–642. - PubMed
-
- Birchmeier W, Behrens J. Cadherin expression in carcinomas: role in the formation of cell junctions and the prevention of invasiveness. Biochim Biophys Acta. 1994;1198:11–26. - PubMed
-
- Bradley, A. 1987. Production and analysis of chimeric mice. In Teratocarcinomas and Embryonic Stem Cells: A Practical Approach. E.J. Robertson, editor. IRL Press, Oxford. 113–151.
-
- Bussemakers MJG, Bokhoven AV, Voller M, Smit FP, Schalken JA. The genes for the calcium-dependent cell adhesion molecules P- and E-cadherin are tandemly arranged in the human genome. Biochem Biophys Res Com. 1994;203:1291–1294. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
