

Metabotropic glutamate receptor-mediated suppression of an inward rectifier current is linked via a cGMP cascade
- PMID: 9364042
- PMCID: PMC6573620
- DOI: 10.1523/JNEUROSCI.17-23-08945.1997
Metabotropic glutamate receptor-mediated suppression of an inward rectifier current is linked via a cGMP cascade
Abstract
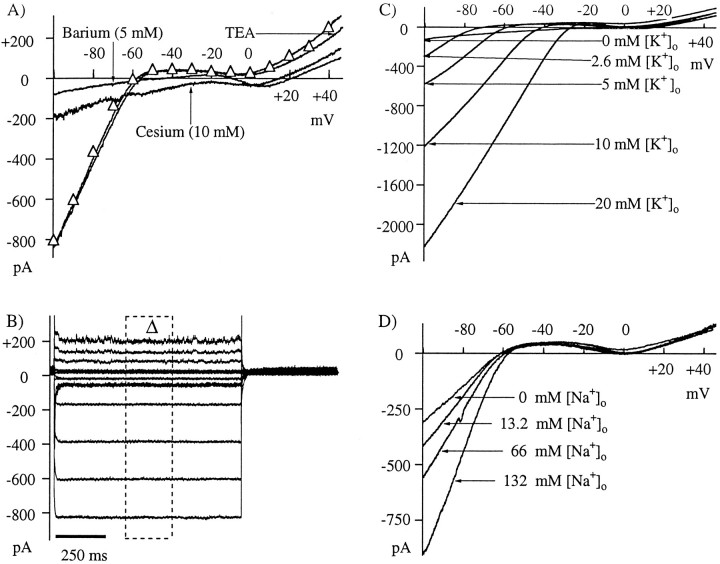
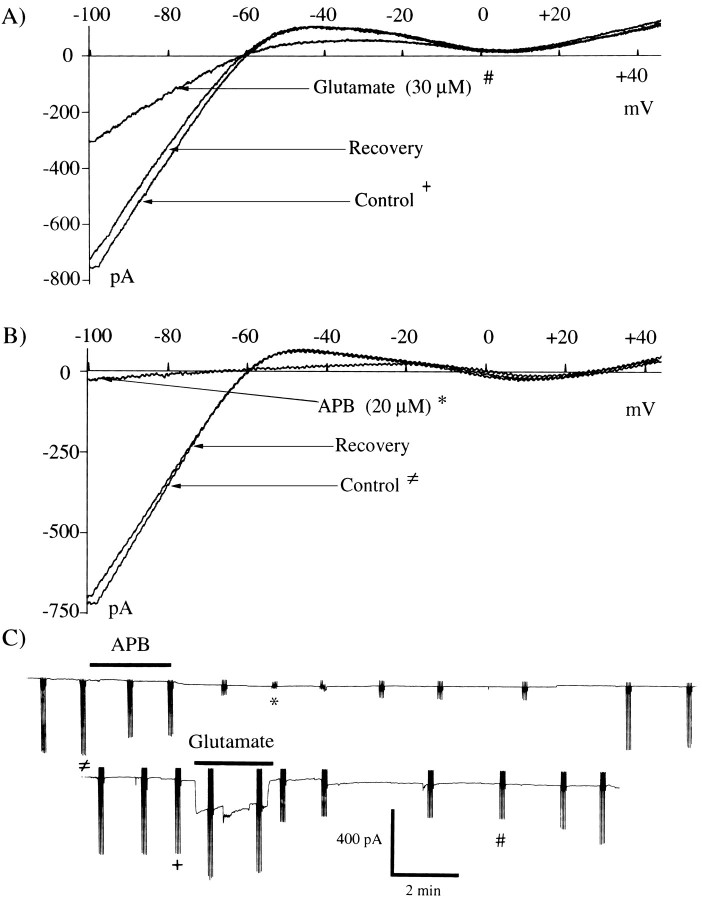
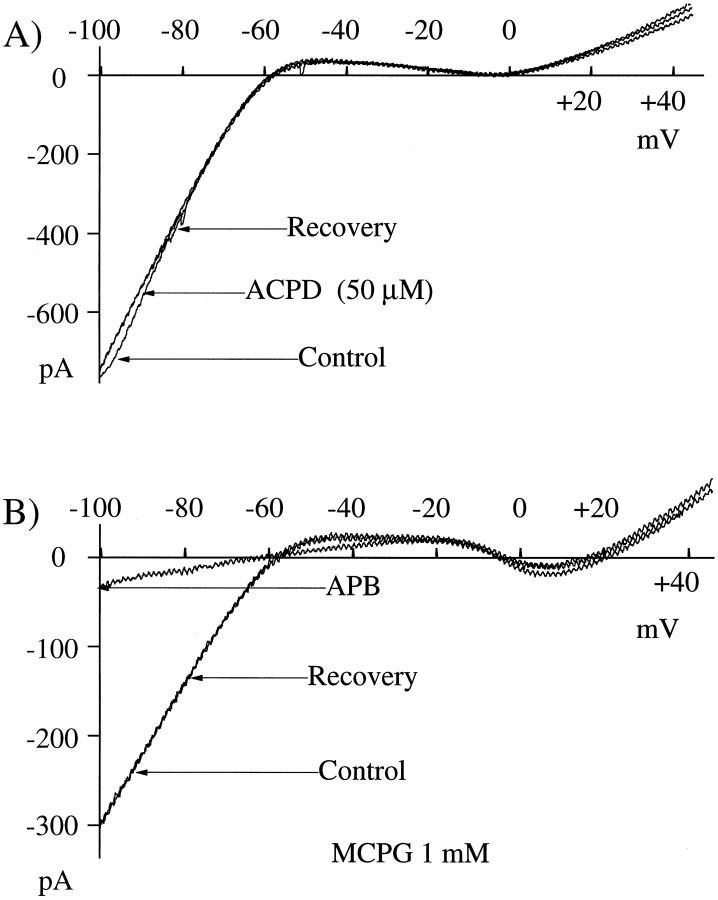
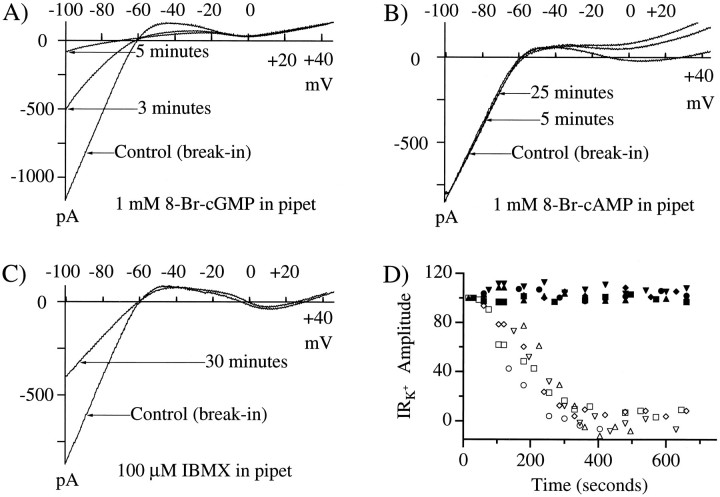
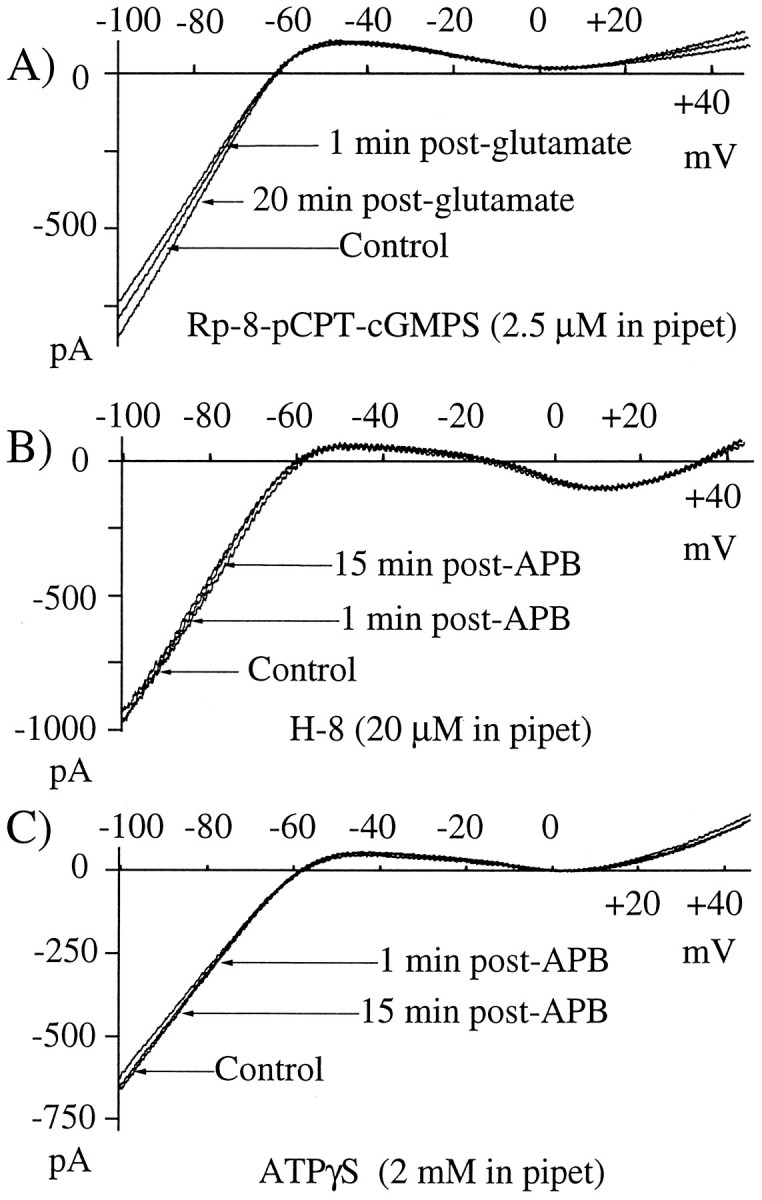
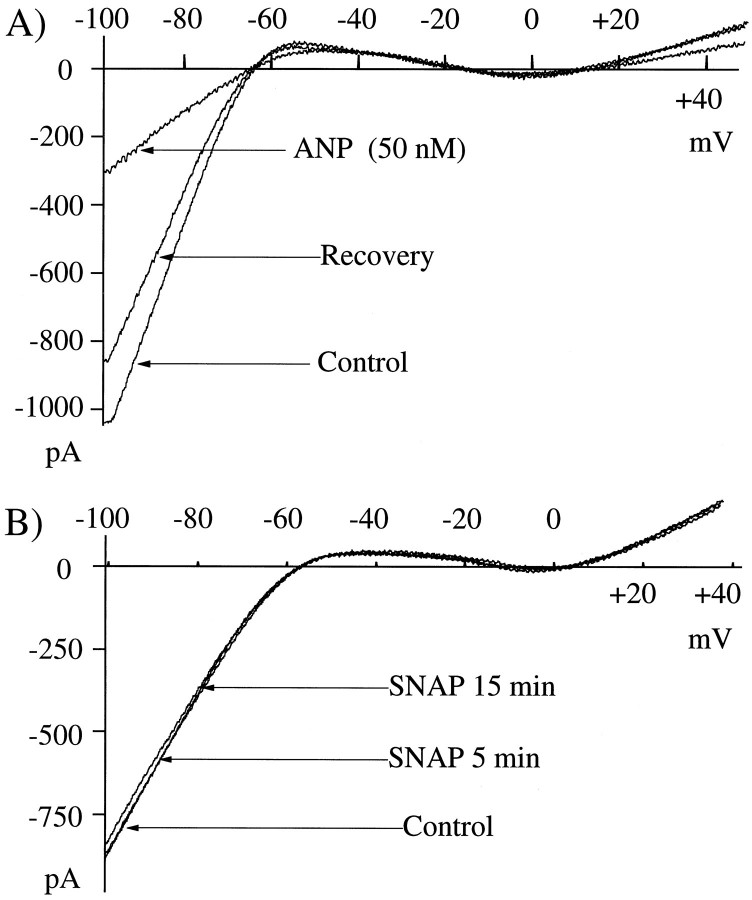
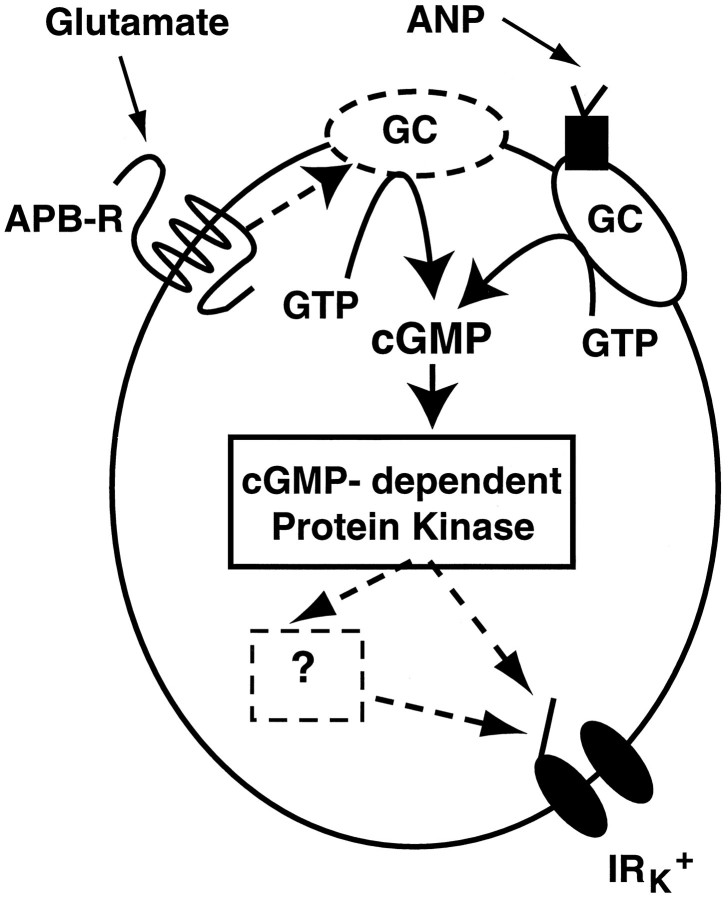
Glutamate, the neurotransmitter released by photoreceptors, excites horizontal cells and OFF-type bipolar cells by activating ionotropic receptors. This study investigated an additional action of glutamate in which it modulates a voltage-gated ion channel in horizontal cells. We find that glutamate and APB (2-amino-4-phosphonobutyrate) produce a delayed and moderately prolonged suppression of an inward rectifier current (IRK+). This effect is proposed to occur via an APB-sensitive metabotropic glutamate receptor (mGluR) because common agonists for the ionotropic or APB-insensitive mGluRs are ineffective and the APB-insensitive receptor antagonist alpha-methyl-4-carboxyphenylglycine (MCPG) does not block the actions of glutamate or APB. 8-Br-cGMP, 1-methyl-3-isobutylxanthine (IBMX), and atrial natriuretic peptide (ANP) but not 8-Br-cAMP mimic the suppression of IRK+. The effects of glutamate and APB are blocked by protein kinase inhibitors including Rp-8-pCPT-cGMPS, H-8, and H-7 as well as by ATPgammaS. We hypothesize that the APB receptor suppresses IRK+ via upregulation of cGMP and subsequent activation of a cGMP-dependent protein kinase. This pathway is likely regulated by an ATP-dependent phosphorylation. This is a novel signaling pathway for mGluRs and indicates that at least two distinct APB-activated pathways exist in the retina. Functionally, this APB receptor-mediated action found in horizontal cells would provide a means by which spatially restricted changes of glutamate, produced by local illumination of photoreceptors, could regulate IRK+ and consequently the response properties of these neurons. This would serve to adapt selectively retinal regions stimulated by small regions of the visual world.
Figures
Similar articles
-
The metabotropic receptor mGluR6 may signal through G(o), but not phosphodiesterase, in retinal bipolar cells.J Neurosci. 1999 Apr 15;19(8):2938-44. doi: 10.1523/JNEUROSCI.19-08-02938.1999. J Neurosci. 1999. PMID: 10191311 Free PMC article.
-
G-protein activation by metabotropic glutamate receptors reduces spike frequency adaptation in neocortical neurons.Neuroscience. 1996 Nov;75(1):123-31. doi: 10.1016/0306-4522(96)00244-8. Neuroscience. 1996. PMID: 8923528
-
Suppression by glutamate of cGMP-activated conductance in retinal bipolar cells.Nature. 1990 Jul 19;346(6281):269-71. doi: 10.1038/346269a0. Nature. 1990. PMID: 1695713
-
Glutamate and Glutamate Receptors in the Vertebrate Retina.2005 May 1 [updated 2007 May 7]. In: Kolb H, Fernandez E, Jones B, Nelson R, editors. Webvision: The Organization of the Retina and Visual System [Internet]. Salt Lake City (UT): University of Utah Health Sciences Center; 1995–. 2005 May 1 [updated 2007 May 7]. In: Kolb H, Fernandez E, Jones B, Nelson R, editors. Webvision: The Organization of the Retina and Visual System [Internet]. Salt Lake City (UT): University of Utah Health Sciences Center; 1995–. PMID: 21413387 Free Books & Documents. Review.
-
Regulation of ON bipolar cell activity.Prog Retin Eye Res. 2008 Jul;27(4):450-63. doi: 10.1016/j.preteyeres.2008.03.003. Epub 2008 Apr 6. Prog Retin Eye Res. 2008. PMID: 18524666 Free PMC article. Review.
Cited by
-
Glutamate modulation of GABA transport in retinal horizontal cells of the skate.J Physiol. 2003 Feb 1;546(Pt 3):717-31. doi: 10.1113/jphysiol.2002.034421. J Physiol. 2003. PMID: 12562999 Free PMC article.
-
Cyclic-nucleotide-gated channels mediate synaptic feedback by nitric oxide.Nature. 1997 Dec 18-25;390(6661):694-8. doi: 10.1038/37803. Nature. 1997. PMID: 9414163 Free PMC article.
-
Metabotropic and ionotropic glutamate receptors regulate calcium channel currents in salamander retinal ganglion cells.J Physiol. 1998 Aug 1;510 ( Pt 3)(Pt 3):815-28. doi: 10.1111/j.1469-7793.1998.815bj.x. J Physiol. 1998. PMID: 9660896 Free PMC article.
-
"mGlu Receptors in the Retina" - WIREs Membrane Transport and Signaling.Wiley Interdiscip Rev Membr Transp Signal. 2012 Sep;1(5):641-653. doi: 10.1002/wmts.43. Wiley Interdiscip Rev Membr Transp Signal. 2012. PMID: 24003403 Free PMC article.
-
Localization of metabotropic glutamate receptors in the outer plexiform layer of the goldfish retina.Cell Tissue Res. 2007 Dec;330(3):389-403. doi: 10.1007/s00441-007-0496-1. Epub 2007 Sep 29. Cell Tissue Res. 2007. PMID: 17906878 Free PMC article.
References
-
- Ahmad I, Leinders-Zufall T, Kocsis JD, Shepherd GM, Zufall F, Barnstable CJ. Retinal ganglion cells express a cGMP-gated cation conductance activatable by nitric oxide donors. Neuron. 1994;12:155–165. - PubMed
-
- Akazawa C, Ohishi H, Nakajima Y, Okamoto N, Shigemoto R, Nakanishi S, Mizuno N. Expression of mRNAs of L-APB-sensitive metabotropic glutamate receptors (mGluR4, mGluR6, mGluR7) in the rat retina. Neurosci Lett. 1994;171:52–54. - PubMed
-
- Akopian A, Witkovsky P. Activation of metabotropic glutamate receptors decreases a high-threshold calcium current in spiking neurons of the Xenopus retina. Vis Neurosci. 1996;13:549–557. - PubMed
-
- Ball AK, Baldridge WH. Background illumination reduces horizontal cell receptive-field size in both normal and 6-hydroxydopamine-lesioned goldfish retinas. Vis Neurosci. 1991;7:441–450. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources