

Transcriptional activity of pannier is regulated negatively by heterodimerization of the GATA DNA-binding domain with a cofactor encoded by the u-shaped gene of Drosophila
- PMID: 9367990
- PMCID: PMC316702
- DOI: 10.1101/gad.11.22.3096
Transcriptional activity of pannier is regulated negatively by heterodimerization of the GATA DNA-binding domain with a cofactor encoded by the u-shaped gene of Drosophila
Abstract
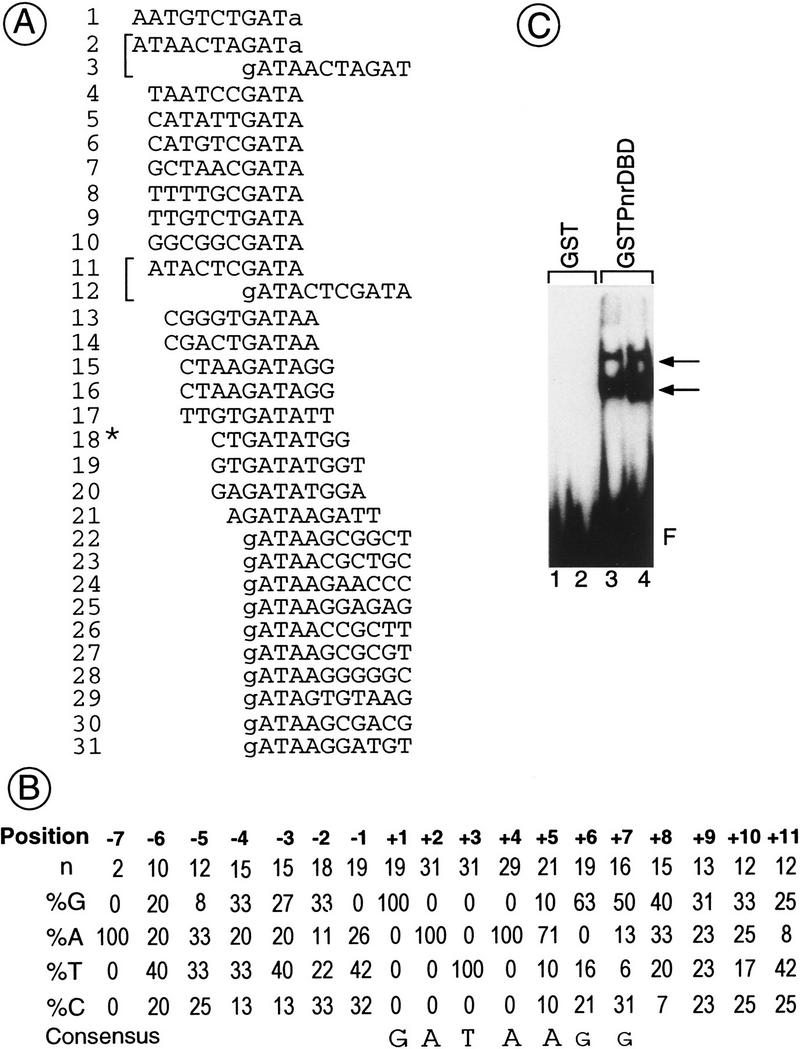
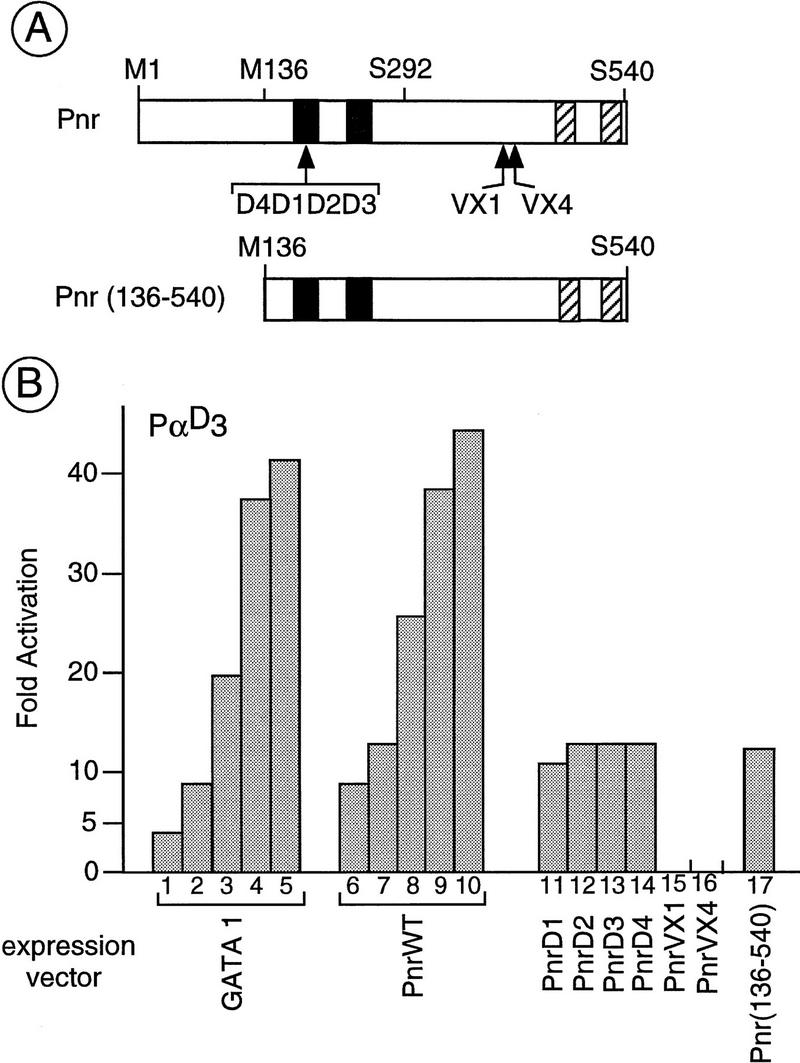
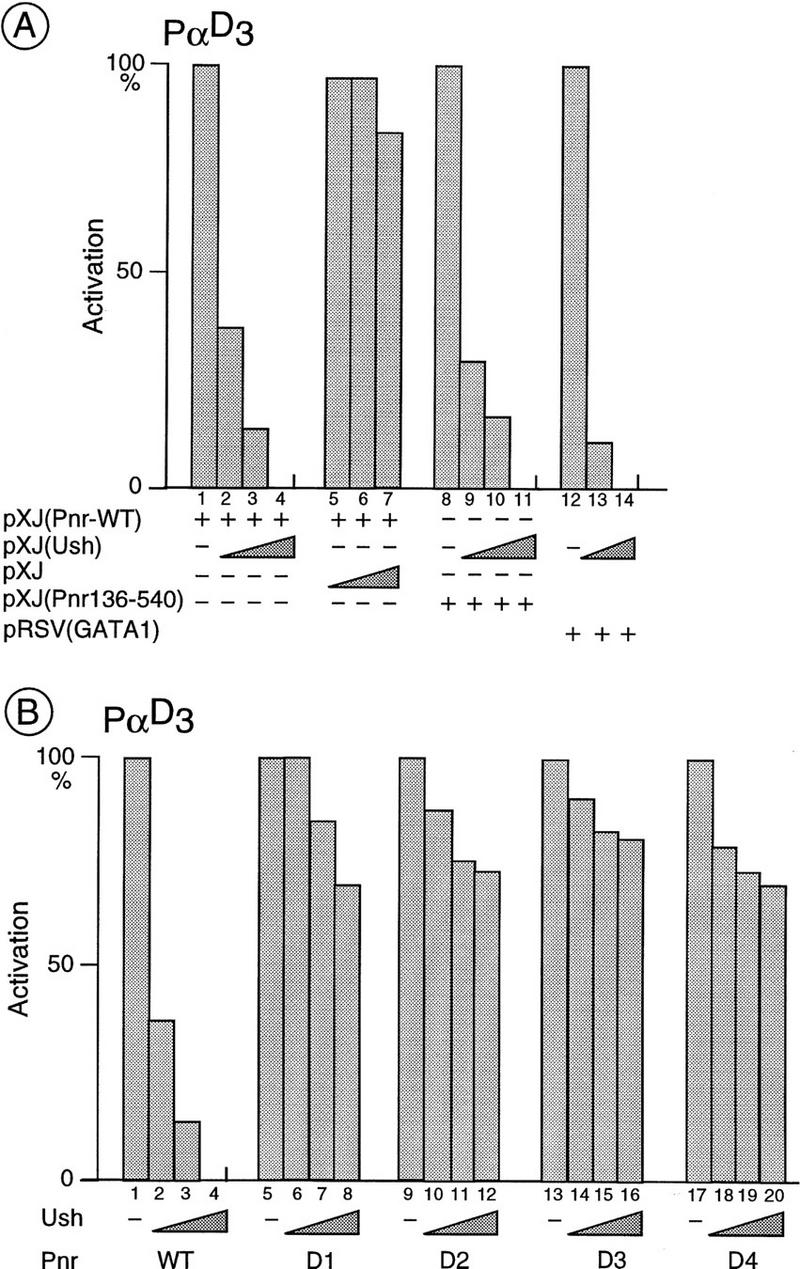
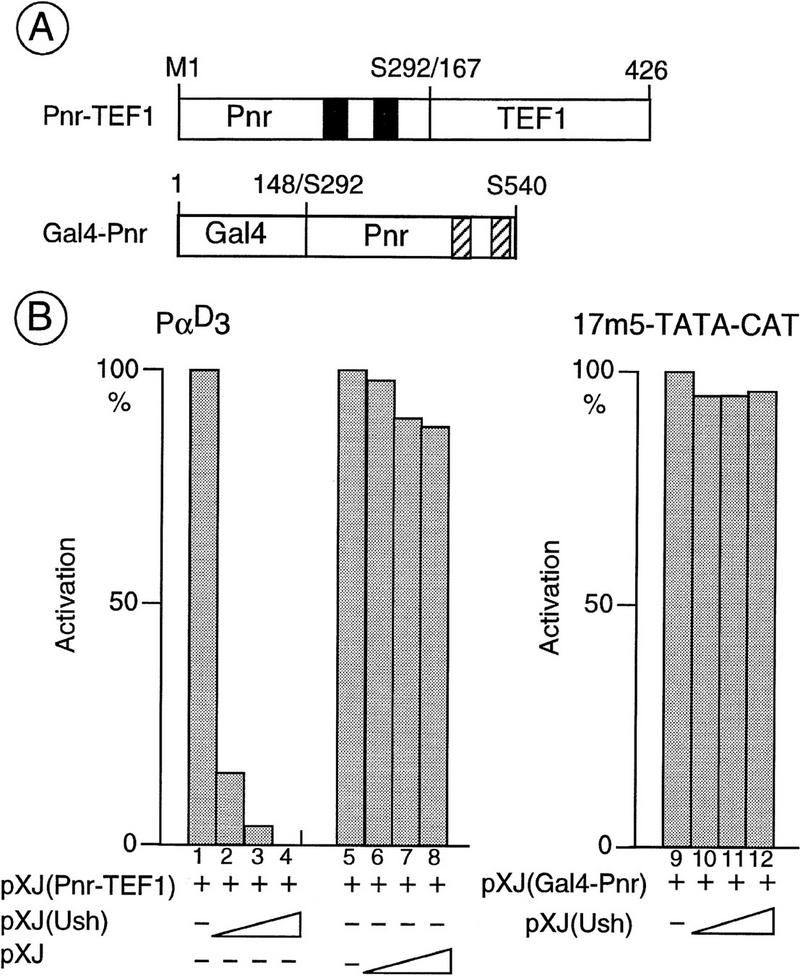
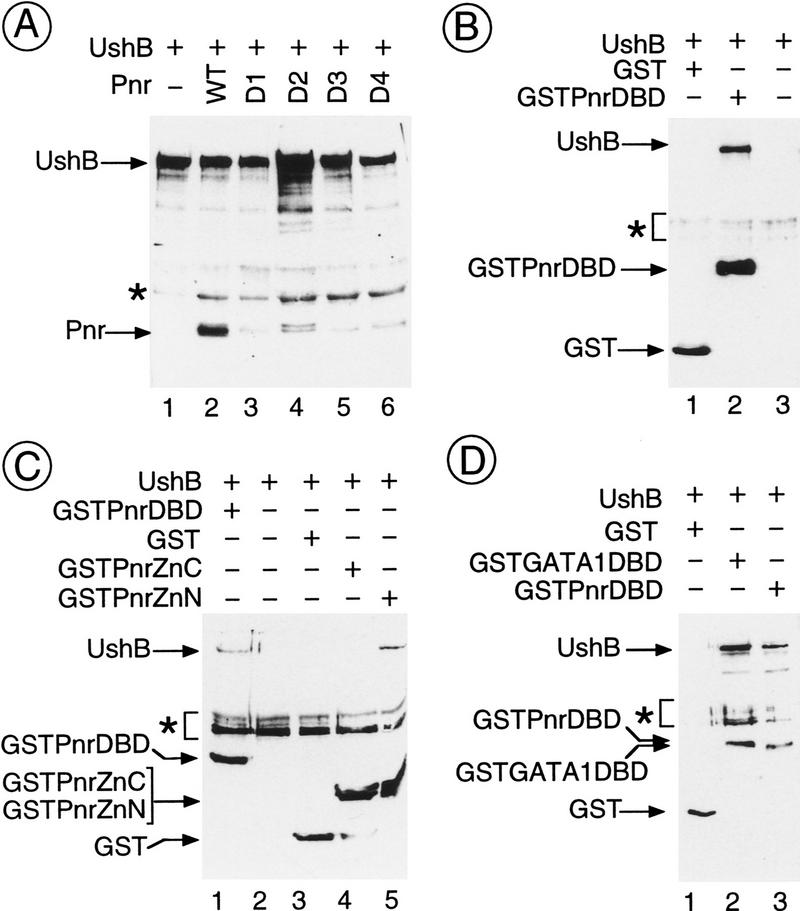
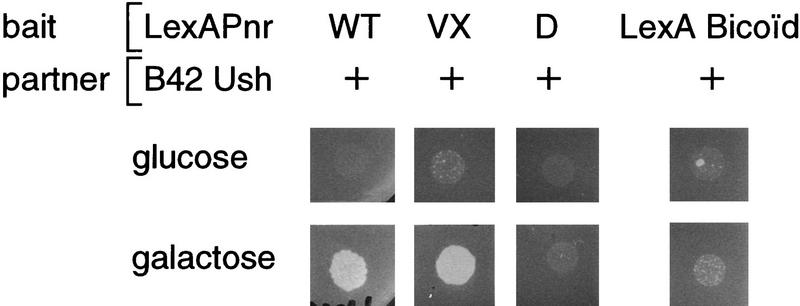
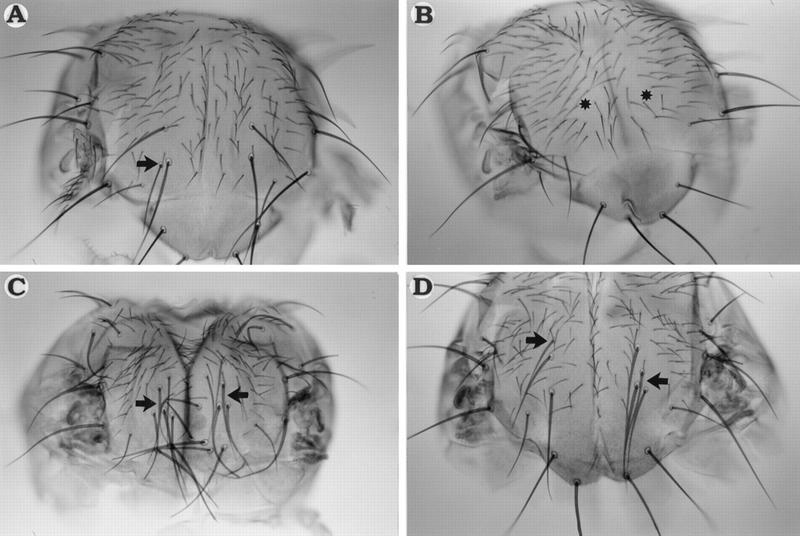
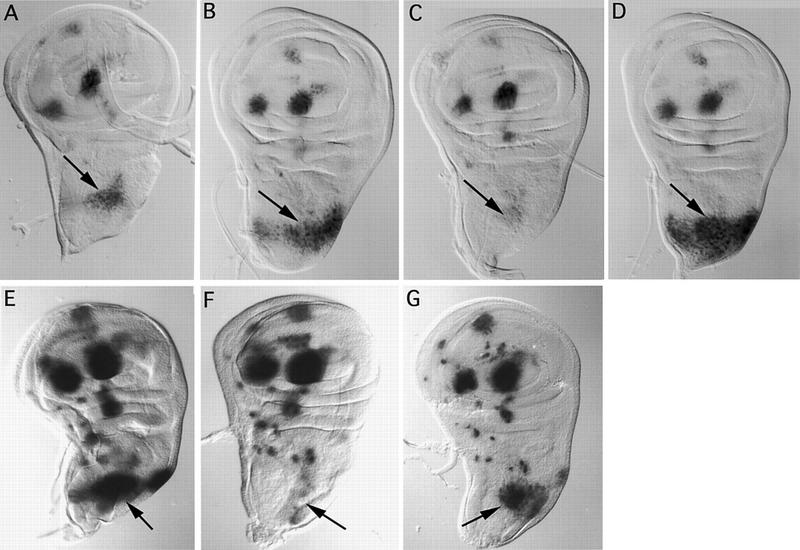
The genes pannier (pnr) and u-shaped (ush) are required for the regulation of achaete-scute during establishment of the bristle pattern in Drosophila. pnr encodes a protein belonging to the GATA family of transcription factors, whereas ush encodes a novel zinc finger protein. Genetic interactions between dominant pnr mutants bearing lesions situated in the amino-terminal zinc finger of the GATA domain and ush mutants have been described. We show here that both wild-type Pannier and the dominant mutant form activate transcription from the heterologous alpha globin promoter when transfected into chicken embryonic fibroblasts. Furthermore, Pnr and Ush are found to heterodimerize through the amino-terminal zinc finger of Pnr and when associated with Ush, the transcriptional activity of Pnr is lost. In contrast, the mutant pnr protein with lesions in this finger associates only poorly with Ush and activates transcription even when cotransfected with Ush. These interactions have been investigated in vivo by overexpression of the mutant and wild-type proteins. The results suggest an antagonistic effect of Ush on Pnr function and reveal a new mode of regulation of GATA factors during development.
Figures
References
-
- Abel T, Michelson AM, Maniatis T. A Drosophila GATA family member that binds to Adh regulatory sequences is expressed in the developing fat body. Development. 1993;119:623–633. - PubMed
-
- Andrews NC, Orkin SH. Transcriptional control of erythropoiesis. Curr Opin Hematol. 1994;1:119–124. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Calleja M, Moreno E, Pelaz S, Morata G. Visualization of gene expression in living adult Drosophila. Science. 1996;274:252–255. - PubMed
-
- Campuzano S, Modolell J. Patterning of the Drosophila nervous system: The achaete-scute gene complex. Trends Genet. 1992;8:202–208. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases