

Two sterol regulatory element-like sequences mediate up-regulation of caveolin gene transcription in response to low density lipoprotein free cholesterol
- PMID: 9380697
- PMCID: PMC23450
- DOI: 10.1073/pnas.94.20.10693
Two sterol regulatory element-like sequences mediate up-regulation of caveolin gene transcription in response to low density lipoprotein free cholesterol
Abstract
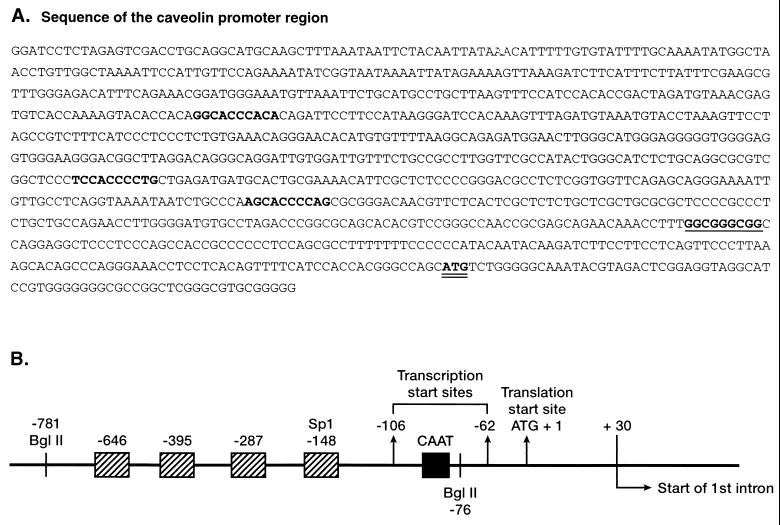
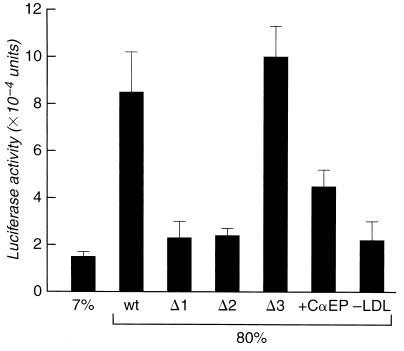
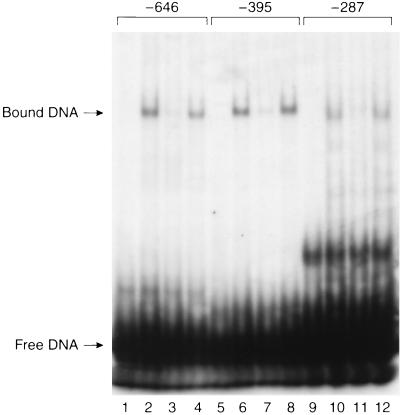
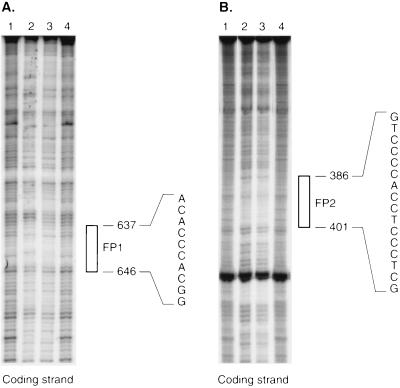
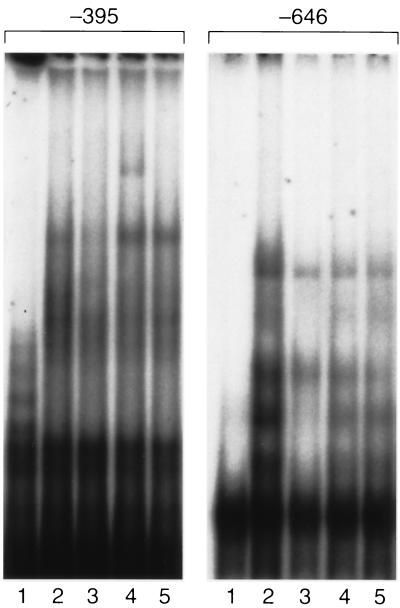
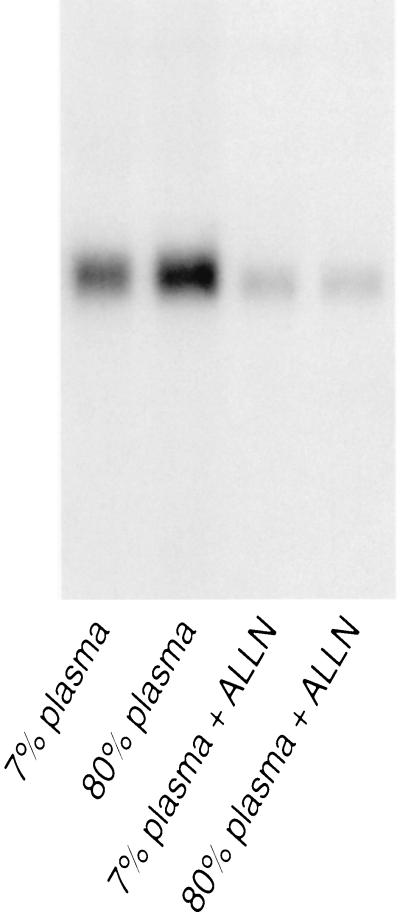
Caveolae form the terminus for a major pathway of intracellular free cholesterol (FC) transport. Caveolin mRNA levels in confluent human skin fibroblasts were up-regulated following increased uptake of low density lipoprotein (LDL) FC. The increase induced by FC was not associated with detectable change in mRNA stability, indicating that caveolin mRNA levels were mediated at the level of gene transcription. A total of 924 bp of 5' flanking region of the caveolin gene were cloned and sequenced. The promoter sequence included three G+C-rich potential sterol regulatory elements (SREs), a CAAT sequence and a Sp1 consensus sequence. Deletional mutagenesis of individual SRE-like sequences indicated that of these two (at -646 and -395 bp) were essential for the increased transcription rates mediated by LDL-FC, whereas the third was inconsequential. Gel shift analysis of protein binding from nuclear extracts to these caveolin promoter DNA sequences, together with DNase I footprinting, confirmed nucleoprotein binding to the SRE-like elements as part of the transcriptional response to LDL-FC. A supershift obtained with antibody to SRE-binding protein 1 (SPEBP-1) indicated that this protein binds at -395 bp. There was no reaction at -395 bp with anti-Sp1 antibody nor with either antibody at -646 bp. The cysteine protease inhibitor N-acetyl-leu-leu-norleucinal (ALLN), which inhibits SREBP catabolism, superinhibited caveolin mRNA levels regardless of LDL-FC. This finding suggests that SREBP inhibits caveolin gene transcription in contrast to its stimulating effect on other promoters. The findings of this study are consistent with the postulated role for caveolin as a regulator of cellular FC homeostasis in quiescent peripheral cells, and the coordinate regulation by SREBP of FC influx and efflux.
Figures
References
-
- Fielding C J, Fielding P E. J Lipid Res. 1997;38:1503–1521. - PubMed
-
- Fielding P E, Fielding C J. Biochemistry. 1995;34:14288–14292. - PubMed
-
- Fielding P E, Fielding C J. Biochemistry. 1996;35:14932–14938. - PubMed
-
- Parton R G, Simons K. Science. 1995;269:1398–1399. - PubMed
-
- Fra A M, Williamson E, Simons K, Parton R G. J Biol Chem. 1994;269:30745–30748. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
