

The unique hetero-oligomeric nature of the subunits in the catalytic cooperativity of the yeast Cct chaperonin complex
- PMID: 9380710
- PMCID: PMC23484
- DOI: 10.1073/pnas.94.20.10780
The unique hetero-oligomeric nature of the subunits in the catalytic cooperativity of the yeast Cct chaperonin complex
Erratum in
- Proc Natl Acad Sci U S A 1997 Dec 23;94(26):14976
Abstract
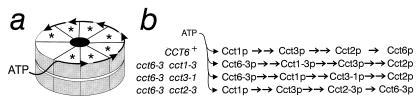
The structural and functional organization of the Cct complex was addressed by genetic analyses of subunit interactions and catalytic cooperativity among five of the eight different essential subunits, Cct1p-Cct8p, in the yeast Saccharomyces cerevisiae. The cct1-1, cct2-3, and cct3-1 alleles, containing mutations at the conserved putative ATP-binding motif, GDGTT, are cold-sensitive, whereas single and multiple replacements of the corresponding motif in Cct6p are well tolerated by the cell. We demonstrated herein that cct6-3 (L19S), but not the parolog cct1-5 (R26I), specifically suppresses the cct1-1, cct2-3, and cct3-1 alleles, and that this suppression can be modulated by mutations in a putative phosphorylation motif, RXS, and the putative ATP-binding pocket of Cct6p. Our results suggest that the Cct ring is comprised of a single hetero-oligomer containing eight subunits of differential functional hierarchy, in which catalytic cooperativity of ATP-binding/hydrolysis takes place in a sequential manner different from the concerted cooperativity proposed for GroEL.
Figures


Similar articles
-
Analysis of mutationally altered forms of the Cct6 subunit of the chaperonin from Saccharomyces cerevisiae.Genetics. 1997 Dec;147(4):1609-33. doi: 10.1093/genetics/147.4.1609. Genetics. 1997. PMID: 9409825 Free PMC article.
-
Physiological effects of unassembled chaperonin Cct subunits in the yeast Saccharomyces cerevisiae.Yeast. 2005 Feb;22(3):219-39. doi: 10.1002/yea.1210. Yeast. 2005. PMID: 15704212
-
Overexpressed ribosomal proteins suppress defective chaperonins in Saccharomyces cerevisiae.FEMS Yeast Res. 2008 Dec;8(8):1236-44. doi: 10.1111/j.1567-1364.2008.00425.x. Epub 2008 Aug 1. FEMS Yeast Res. 2008. PMID: 18680526
-
Cystosolic chaperonin subunits have a conserved ATPase domain but diverged polypeptide-binding domains.Trends Biochem Sci. 1994 Dec;19(12):543-8. doi: 10.1016/0968-0004(94)90058-2. Trends Biochem Sci. 1994. PMID: 7846767 Review.
-
Review: the Cct eukaryotic chaperonin subunits of Saccharomyces cerevisiae and other yeasts.Yeast. 1996 May;12(6):523-9. doi: 10.1002/(SICI)1097-0061(199605)12:6%3C523::AID-YEA962%3E3.0.CO;2-C. Yeast. 1996. PMID: 8771707 Review.
Cited by
-
A structural vista of phosducin-like PhLP2A-chaperonin TRiC cooperation during the ATP-driven folding cycle.Nat Commun. 2024 Feb 2;15(1):1007. doi: 10.1038/s41467-024-45242-x. Nat Commun. 2024. PMID: 38307855 Free PMC article.
-
Folding of large multidomain proteins by partial encapsulation in the chaperonin TRiC/CCT.Proc Natl Acad Sci U S A. 2012 Dec 26;109(52):21208-15. doi: 10.1073/pnas.1218836109. Epub 2012 Nov 28. Proc Natl Acad Sci U S A. 2012. PMID: 23197838 Free PMC article.
-
Functional Subunits of Eukaryotic Chaperonin CCT/TRiC in Protein Folding.J Amino Acids. 2011;2011:843206. doi: 10.4061/2011/843206. Epub 2011 Jul 2. J Amino Acids. 2011. PMID: 22312474 Free PMC article.
-
Sequential action of ATP-dependent subunit conformational change and interaction between helical protrusions in the closure of the built-in lid of group II chaperonins.J Biol Chem. 2008 Dec 12;283(50):34773-84. doi: 10.1074/jbc.M805303200. Epub 2008 Oct 13. J Biol Chem. 2008. PMID: 18854314 Free PMC article.
-
The inter-ring arrangement of the cytosolic chaperonin CCT.EMBO Rep. 2007 Mar;8(3):252-7. doi: 10.1038/sj.embor.7400894. Epub 2007 Feb 16. EMBO Rep. 2007. PMID: 17304242 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
