

The mechanism of cancer-mediated conversion of plasminogen to the angiogenesis inhibitor angiostatin
- PMID: 9380726
- PMCID: PMC23512
- DOI: 10.1073/pnas.94.20.10868
The mechanism of cancer-mediated conversion of plasminogen to the angiogenesis inhibitor angiostatin
Abstract
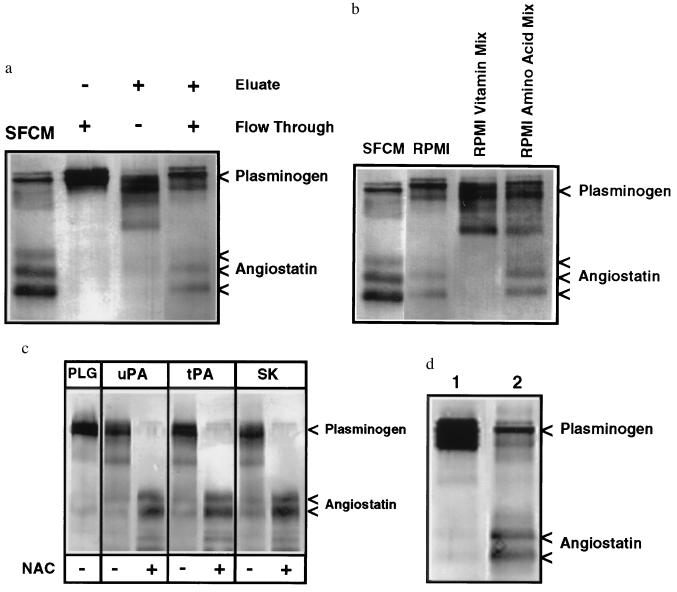
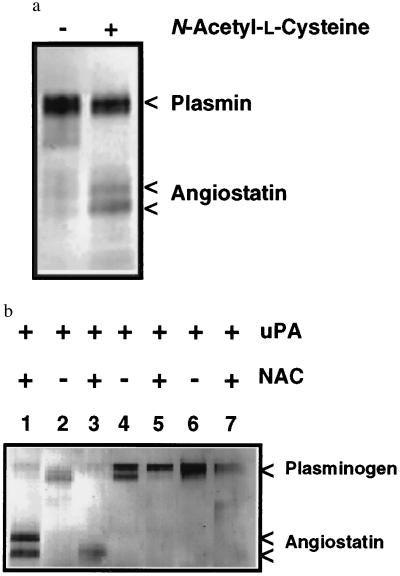
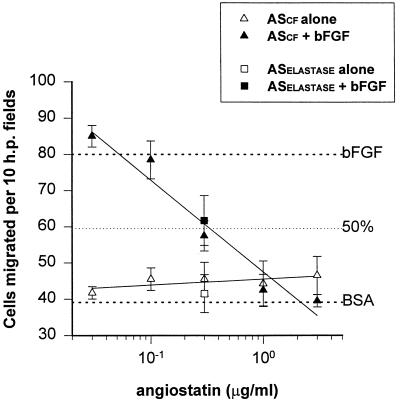
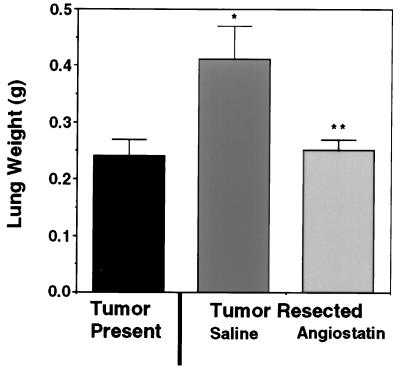
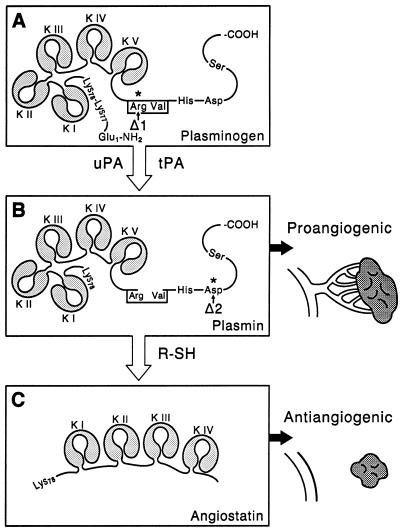
Angiostatin, a potent naturally occurring inhibitor of angiogenesis and growth of tumor metastases, is generated by cancer-mediated proteolysis of plasminogen. Human prostate carcinoma cells (PC-3) release enzymatic activity that converts plasminogen to angiostatin. We have now identified two components released by PC-3 cells, urokinase (uPA) and free sulfhydryl donors (FSDs), that are sufficient for angiostatin generation. Furthermore, in a defined cell-free system, plasminogen activators [uPA, tissue-type plasminogen activator (tPA), or streptokinase], in combination with one of a series of FSDs (N-acetyl-L-cysteine, D-penicillamine, captopril, L-cysteine, or reduced glutathione] generate angiostatin from plasminogen. An essential role of plasmin catalytic activity for angiostatin generation was identified by using recombinant mutant plasminogens as substrates. The wild-type recombinant plasminogen was converted to angiostatin in the setting of uPA/FSD; however, a plasminogen activation site mutant and a catalytically inactive mutant failed to generate angiostatin. Cell-free derived angiostatin inhibited angiogenesis in vitro and in vivo and suppressed the growth of Lewis lung carcinoma metastases. These findings define a direct mechanism for cancer-cell-mediated angiostatin generation and permit large-scale production of bioactive angiostatin for investigation and potential therapeutic application.
Figures
References
-
- Folkman J. J Natl Cancer Inst. 1990;82:4–6. - PubMed
-
- Folkman J, Watson K, Ingber D, Hanahan D. Nature (London) 1989;339:58–61. - PubMed
-
- Weidner N, Semple J P, Welch W R, Folkman J. N Engl J Med. 1991;324:1–8. - PubMed
-
- Fotsis T, Zhang Y, Pepper M S, Adlercreutz H, Montesano R, Nawroth P P, Schweigerer L. Nature (London) 1994;368:237–239. - PubMed
-
- Kim K J, Li B, Winer J, Armanini M, Gillett N, Phillips H S, Ferrara N. Nature (London) 1993;362:841–844. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
