

The interaction of the general anesthetic etomidate with the gamma-aminobutyric acid type A receptor is influenced by a single amino acid
- PMID: 9380754
- PMCID: PMC23576
- DOI: 10.1073/pnas.94.20.11031
The interaction of the general anesthetic etomidate with the gamma-aminobutyric acid type A receptor is influenced by a single amino acid
Abstract
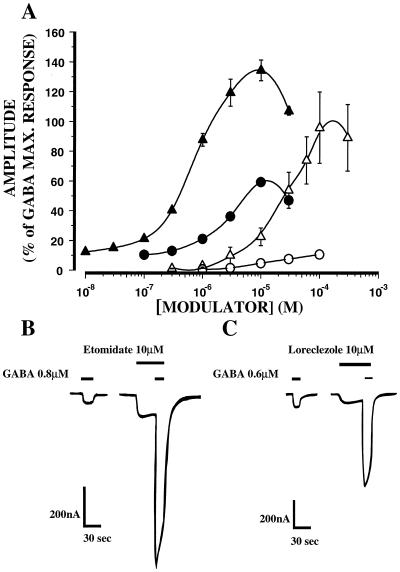
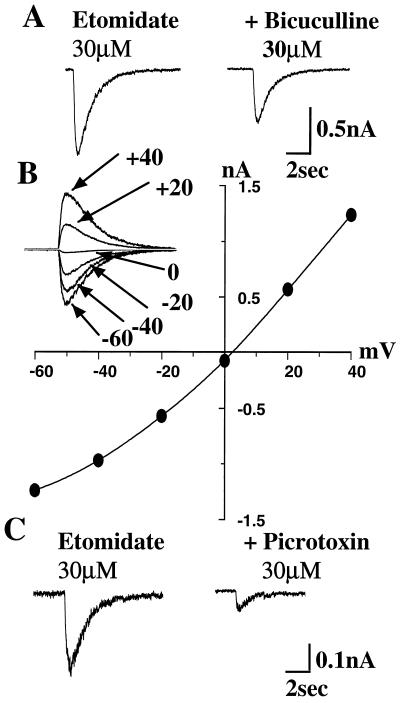
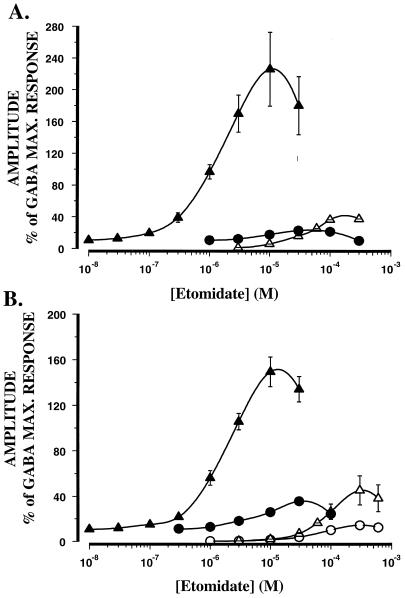
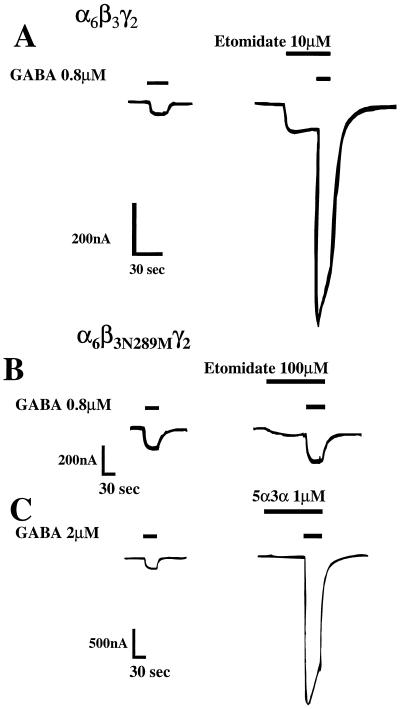
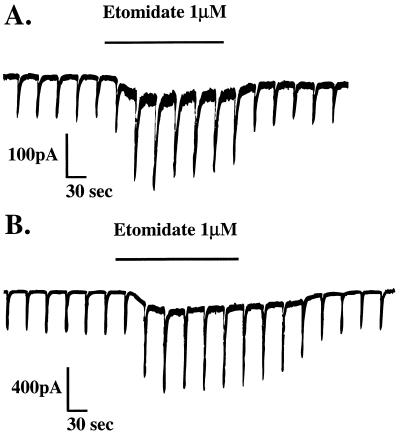
The gamma-aminobutyric acid type A (GABAA) receptor is a transmitter-gated ion channel mediating the majority of fast inhibitory synaptic transmission within the brain. The receptor is a pentameric assembly of subunits drawn from multiple classes (alpha1-6, beta1-3, gamma1-3, delta1, and epsilon1). Positive allosteric modulation of GABAA receptor activity by general anesthetics represents one logical mechanism for central nervous system depression. The ability of the intravenous general anesthetic etomidate to modulate and activate GABAA receptors is uniquely dependent upon the beta subunit subtype present within the receptor. Receptors containing beta2- or beta3-, but not beta1 subunits, are highly sensitive to the agent. Here, chimeric beta1/beta2 subunits coexpressed in Xenopus laevis oocytes with human alpha6 and gamma2 subunits identified a region distal to the extracellular N-terminal domain as a determinant of the selectivity of etomidate. The mutation of an amino acid (Asn-289) present within the channel domain of the beta3 subunit to Ser (the homologous residue in beta1), strongly suppressed the GABA-modulatory and GABA-mimetic effects of etomidate. The replacement of the beta1 subunit Ser-290 by Asn produced the converse effect. When applied intracellularly to mouse L(tk-) cells stably expressing the alpha6beta3gamma2 subunit combination, etomidate was inert. Hence, the effects of a clinically utilized general anesthetic upon a physiologically relevant target protein are dramatically influenced by a single amino acid. Together with the lack of effect of intracellular etomidate, the data argue against a unitary, lipid-based theory of anesthesia.
Figures
References
-
- Rushman G B, Davies N J H, Atkinson R S. A Short History of Anaesthesia. Oxford: Butterworth–Heinemann; 1996. pp. 1–218.
-
- Franks N P, Lieb W R. Nature (London) 1994;367:607–614. - PubMed
-
- Franks N P, Lieb W R. Trends Pharmacol Sci. 1987;8:169–174.
-
- Raines D E, Miller K W. Anesth Analg. 1994;79:1031–1033. - PubMed
-
- Liu J, Laster M J, Taheri S, Eger E I, II, Chortkoff B, Halsey M J. Anesth Analg. 1994;79:1049–1055. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
