

Qualitative regulation of B cell antigen receptor signaling by CD19: selective requirement for PI3-kinase activation, inositol-1,4,5-trisphosphate production and Ca2+ mobilization
- PMID: 9382888
- PMCID: PMC2199152
- DOI: 10.1084/jem.186.11.1897
Qualitative regulation of B cell antigen receptor signaling by CD19: selective requirement for PI3-kinase activation, inositol-1,4,5-trisphosphate production and Ca2+ mobilization
Abstract
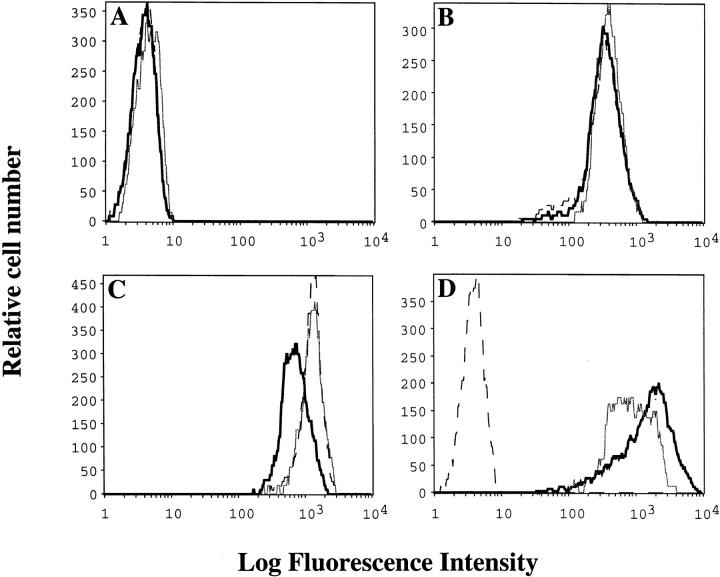
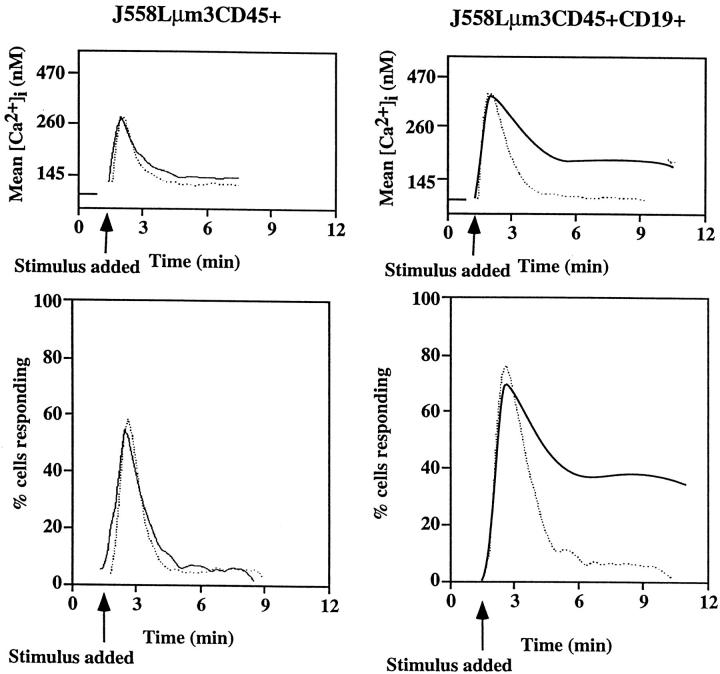
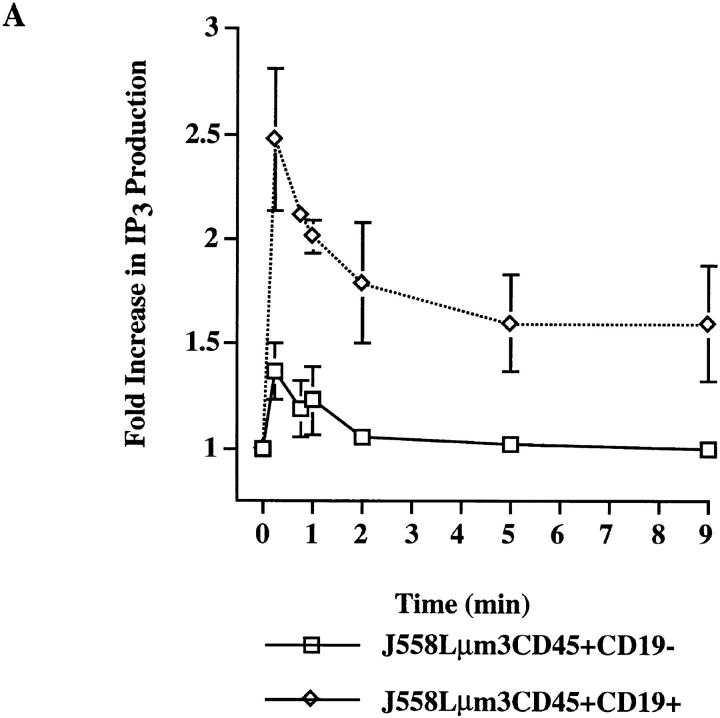
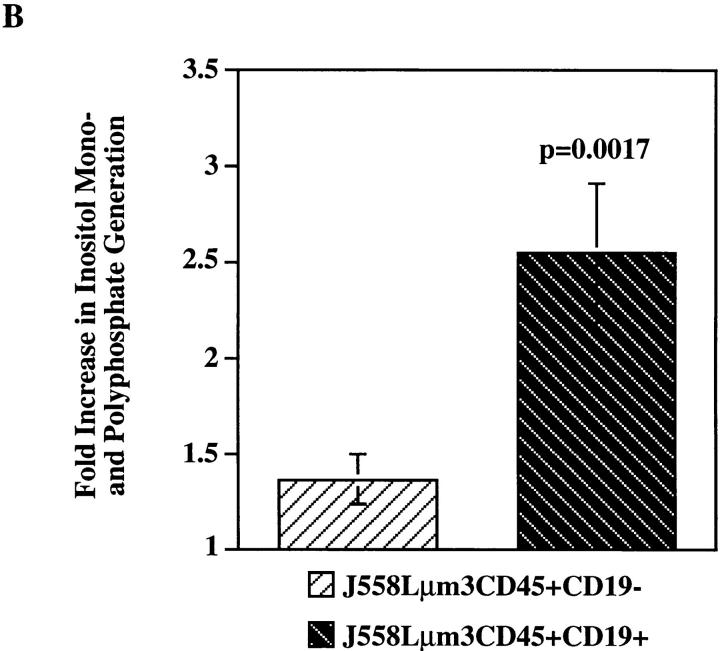
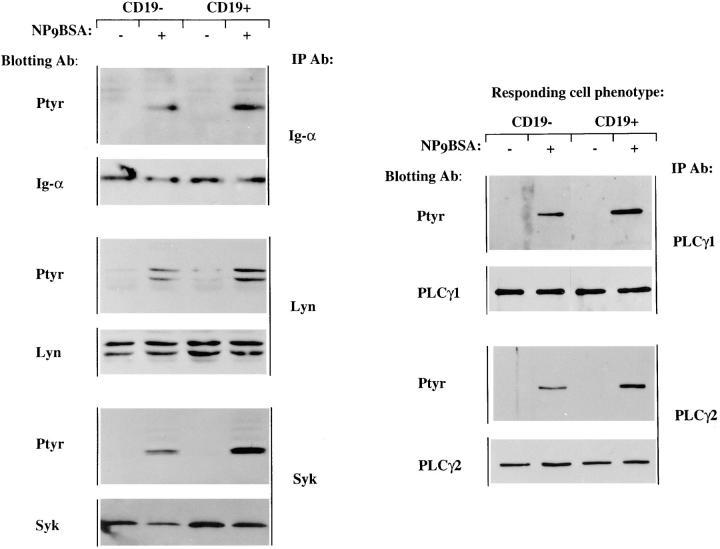
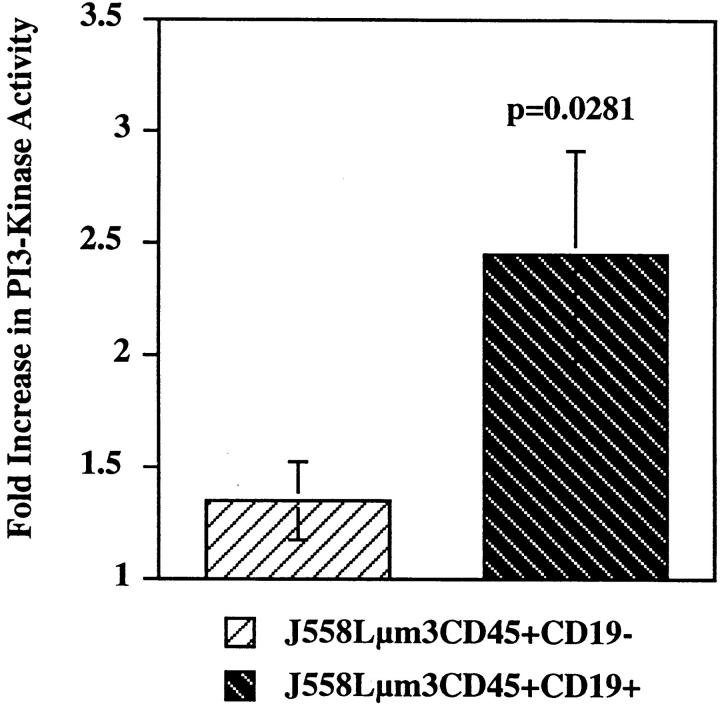
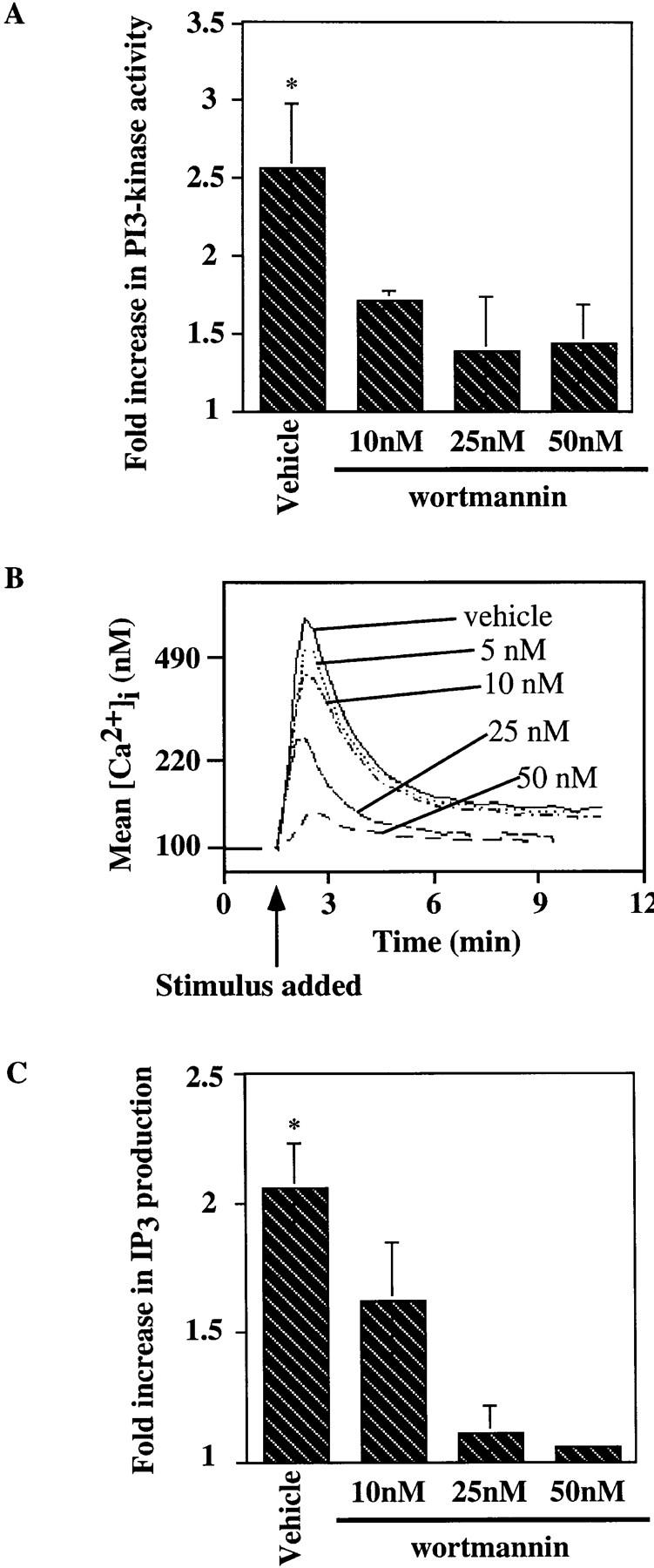
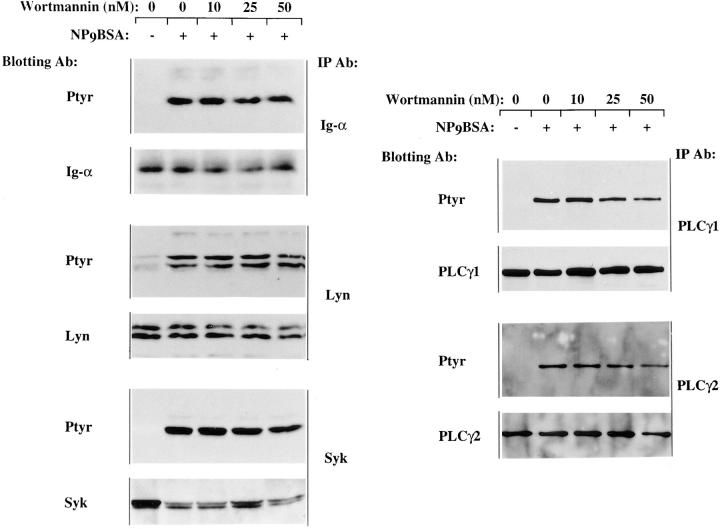
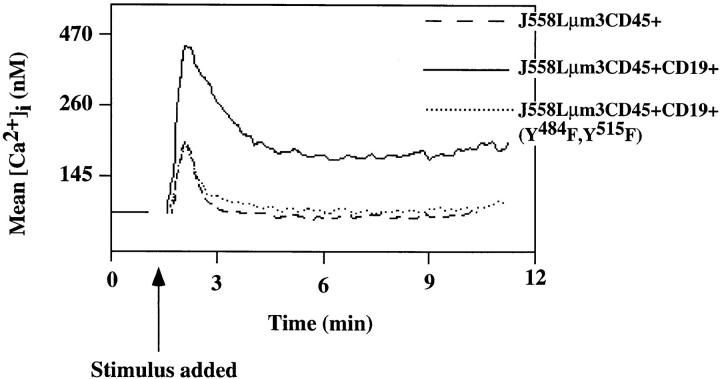
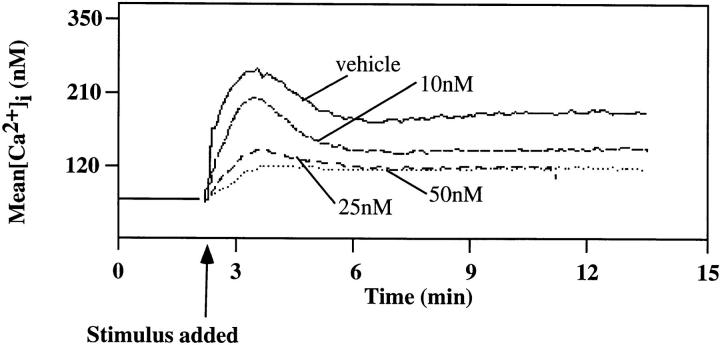
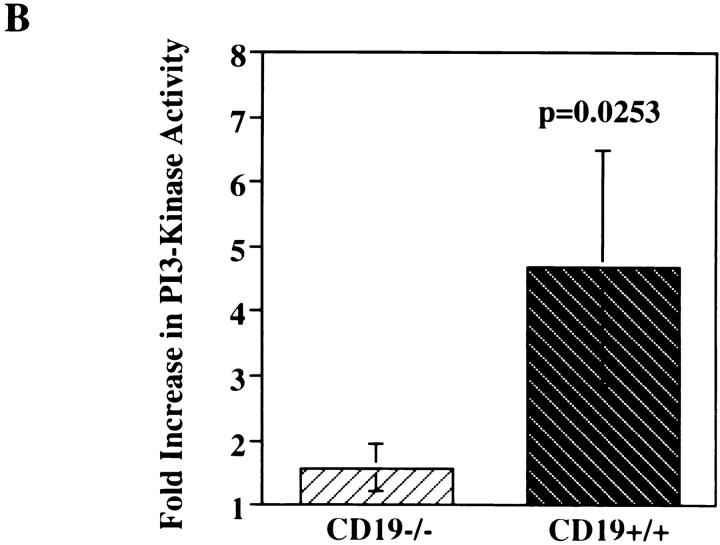
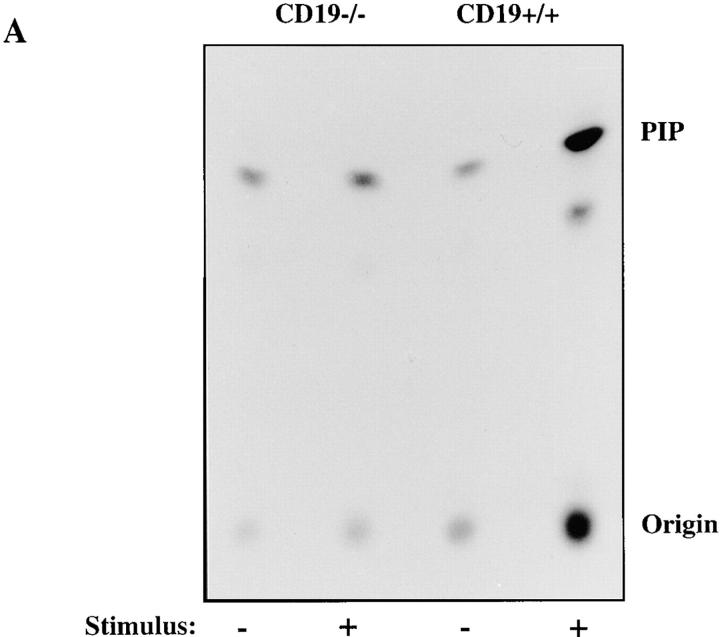
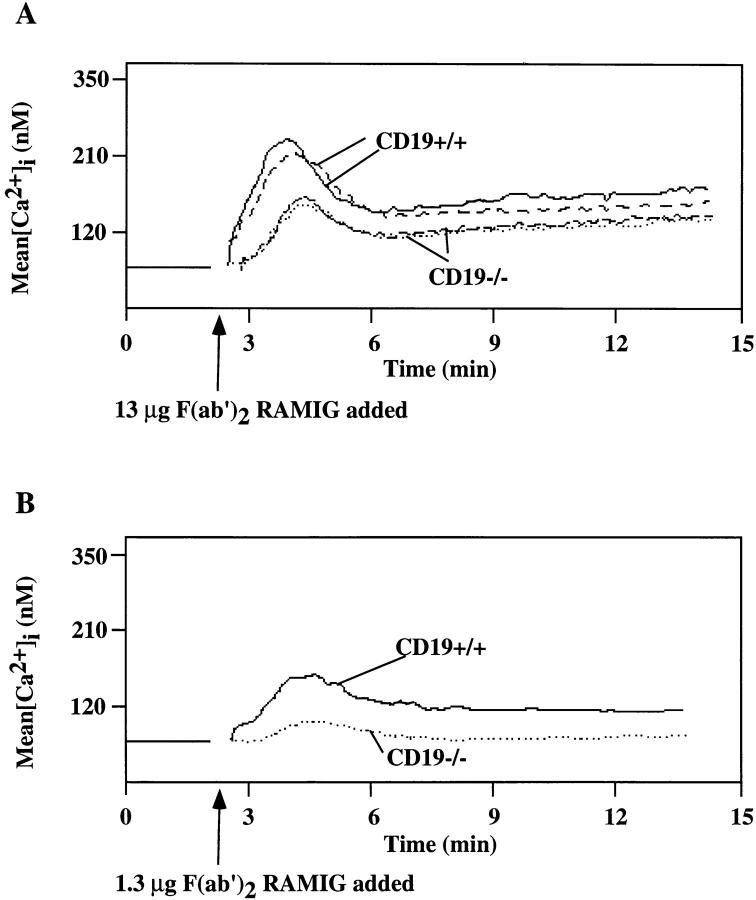
Genetic ablation of the B cell surface glycoprotein CD19 severely impairs the humoral immune response. This requirement is thought to reflect a critical role of CD19 in signal transduction that occurs upon antigen C3dg coligation of antigen receptors with CD19 containing type 2 complement receptors (CR2). Here we show that CD19 plays a key accessory role in B cell antigen receptor signaling independent of CR2 coligation and define molecular circuitry by which this function is mediated. While CD19 is not required for antigen-mediated activation of receptor proximal tyrosines kinases, it is critical for activation of phosphatidylinositol 3-kinase (PI3-kinase). PI3-Kinase activation is dependent on phosphorylation of CD19 Y484 and Y515. Antigen-induced CD19-dependent PI3-kinase activation is required for normal phosphoinositide hydrolysis and Ca2+ mobilization responses. Thus, CD19 functions as a B cell antigen receptor accessory molecule that modifies antigen receptor signaling in a qualitative manner.
Figures
References
-
- Fearon DT, Carter RH. The CD19/CR2/ TAPA-1 complex of B lymphocytes: linking natural to acquired immunity. Ann Rev Immunol. 1995;13:127–149. - PubMed
-
- Carter RH, Doody GM, Bolen JB, Fearon DT. Membrane IgM-induced tyrosine phosphorylation of CD19 requires a CD19 domain that mediates association with components of the B cell antigen receptor complex. J Immunol. 1997;158:3062–3069. - PubMed
-
- Tedder TF, Isaacs CM. Isolation of cDNAs encoding the CD19 antigen of human and mouse B lymphocytes. A new member of the immunoglobulin superfamily. J Immunol. 1989;143:712–717. - PubMed
-
- Roifman CM, Ke S. CD19 is a substrate of the antigen receptor-associated protein tyrosine kinase in human B cells. Biochem Biophys Res Commun. 1993;194:222–225. - PubMed
-
- Uckun FM, Burkhardt AL, Jarvis L, Jun X, Stealey B, Dibirdik I, Myers DE, Tuel AL, Bolen JB. Signal transduction through the CD19 receptor during discrete developmental stages of human B-cell ontogeny. J Biol Chem. 1993;268:21172–21184. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
