

A novel cytosolic regulator, Pianissimo, is required for chemoattractant receptor and G protein-mediated activation of the 12 transmembrane domain adenylyl cyclase in Dictyostelium
- PMID: 9389653
- PMCID: PMC316743
- DOI: 10.1101/gad.11.23.3218
A novel cytosolic regulator, Pianissimo, is required for chemoattractant receptor and G protein-mediated activation of the 12 transmembrane domain adenylyl cyclase in Dictyostelium
Abstract
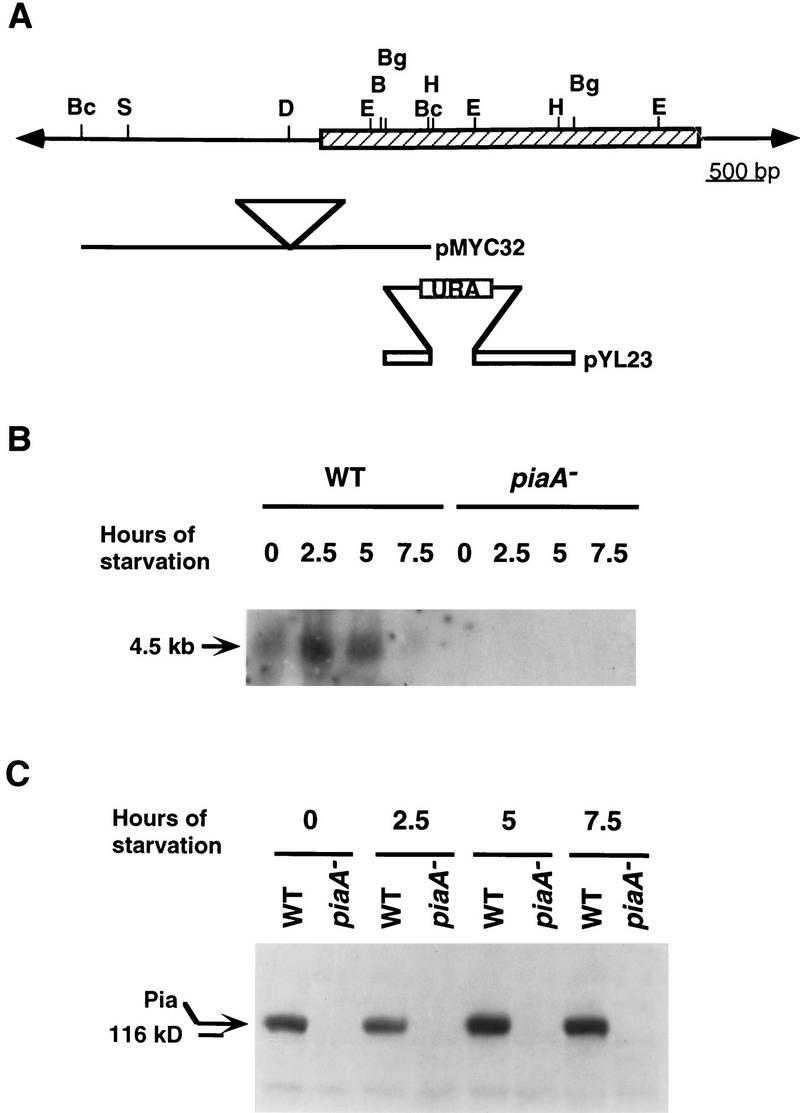
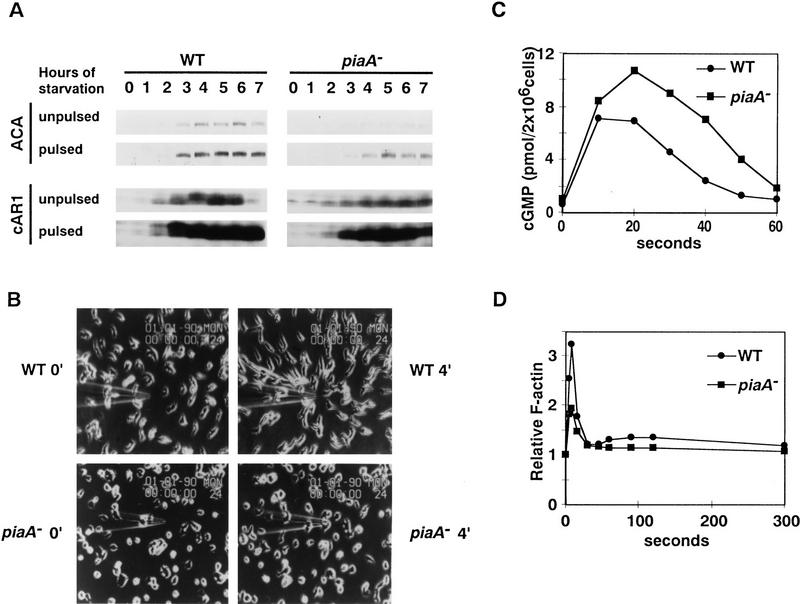
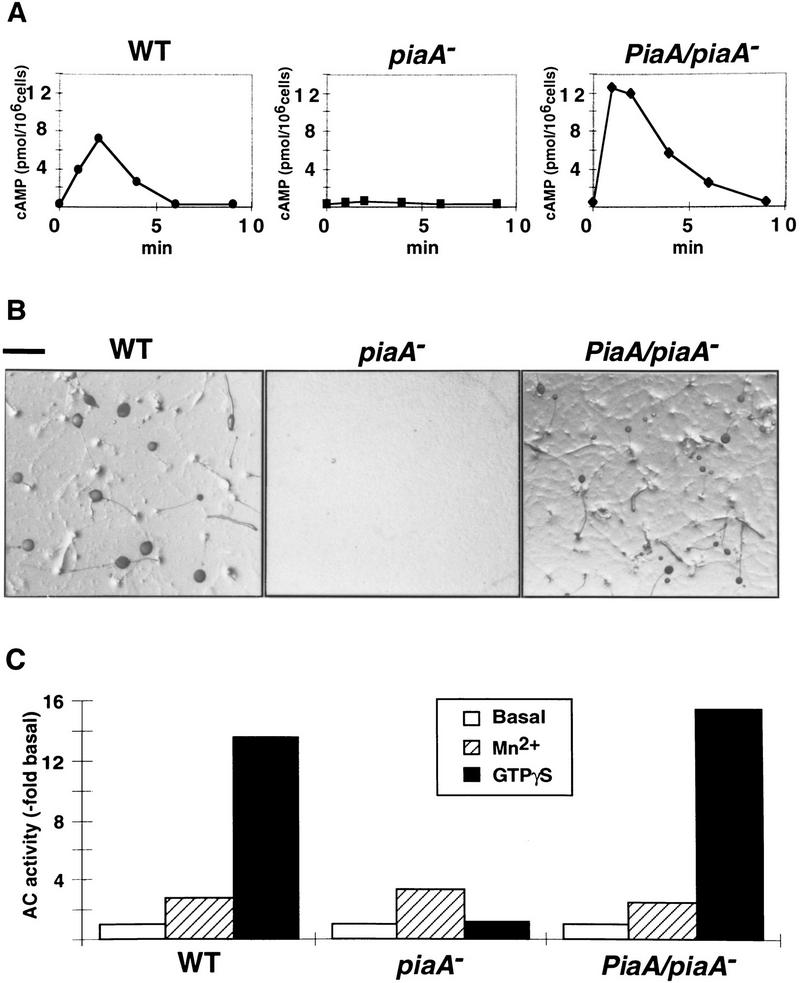
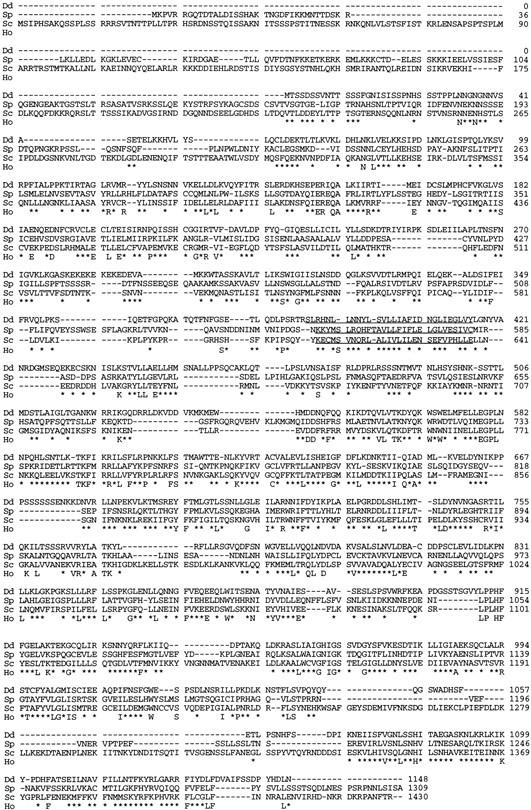
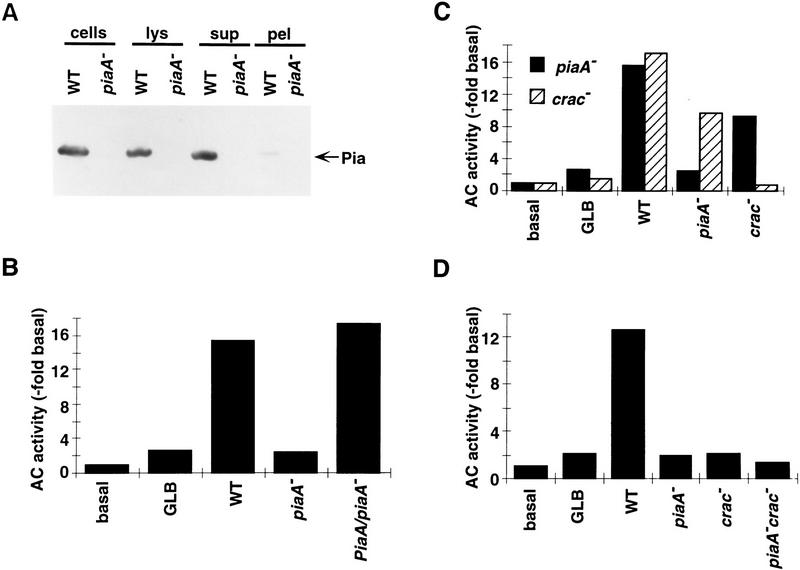
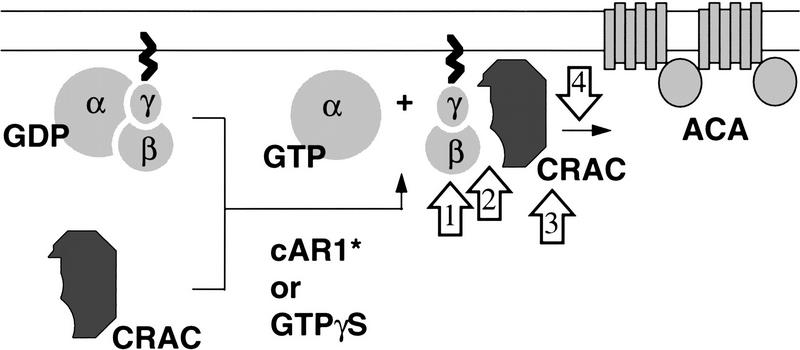
Genetic analysis was applied to identify novel genes involved in G protein-linked pathways controlling development. Using restriction enzyme-mediated integration (REMI), we have identified a new gene, Pianissimo (PiaA), involved in cAMP signaling in Dictyostelium discoideum. PiaA encodes a 130-kD cytosolic protein required for chemoattractant receptor and G protein-mediated activation of the 12 transmembrane domain adenylyl cyclase. In piaA- null mutants, neither chemoattractant stimulation of intact cells nor GTPgammaS treatment of lysates activates the enzyme; constitutive expression of PiaA reverses these defects. Cytosols of wild-type cells that contain Pia protein reconstitute the GTPgammaS stimulation of adenylyl cyclase activity in piaA- lysates, indicating that Pia is directly involved in the activation. Pia and CRAC, a previously identified cytosolic regulator, are both essential for activation of the enzyme as lysates of crac- piaA- double mutants require both proteins for reconstitution. Homologs of PiaA are found in Saccharomyces cerevisiae and Schizosaccaromyces pombe; disruption of the S. cerevisiae homolog results in lethality. We propose that homologs of Pia and similar modes of regulation of these ubiquitous G protein-linked pathways are likely to exist in higher eukaryotes.
Figures
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Broek D, Samiy N, Fasano O, Fujiyama A, Tamanoi F, Northup J, Wigler M. Differential activation of yeast adenylate cyclase by wild-type and mutant RAS proteins. Cell. 1985;41:763–760. - PubMed
-
- Caterina MJ, Devreotes PN. Molecular insights into eukaryotic chemotaxis. FASEB J. 1991;5:3078–3085. - PubMed
-
- Chen J, DeVivo M, Dingus J, Harry A, Li J, Sui J, Carty DJ, Blank JL, Exton JH, Stoffel RH, Inglese J, Lefkowitz RJ, Logothetis DE, Hildebrandt JD, Iyengar R. A region of adenylyl cyclase 2 critical for regulation by G protein βγ subunits. Science. 1995;268:1166–1169. - PubMed
-
- Chen M-Y, Devreotes PN, Gundersen RE. Serine 113 is the site of receptor-mediated phosphorylation of the Dictyostelium G protein α-subunit Gα2. J Biol Chem. 1994;269:20925–20930. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases