

Chaperone-supervised conversion of prion protein to its protease-resistant form
- PMID: 9391131
- PMCID: PMC28411
- DOI: 10.1073/pnas.94.25.13938
Chaperone-supervised conversion of prion protein to its protease-resistant form
Abstract
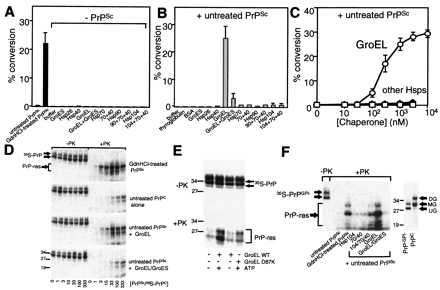
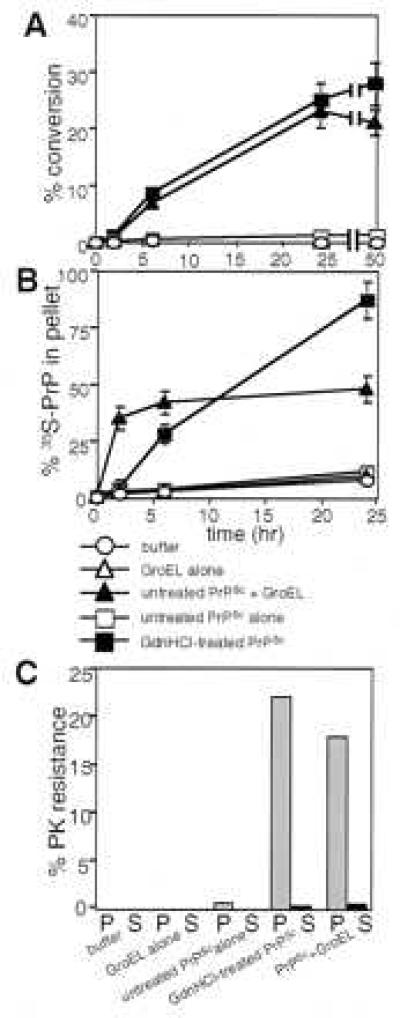
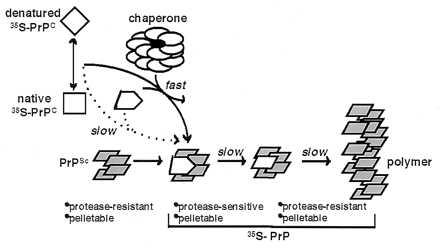
Transmissible spongiform encephalopathies (TSEs) are lethal, infectious disorders of the mammalian nervous system. A TSE hallmark is the conversion of the cellular protein PrPC to disease-associated PrPSc (named for scrapie, the first known TSE). PrPC is protease-sensitive, monomeric, detergent soluble, and primarily alpha-helical; PrPSc is protease-resistant, polymerized, detergent insoluble, and rich in beta-sheet. The "protein-only" hypothesis posits that PrPSc is the infectious TSE agent that directly converts host-encoded PrPC to fresh PrPSc, harming neurons and creating new agents of infection. To gain insight on the conformational transitions of PrP, we tested the ability of several protein chaperones, which supervise the conformational transitions of proteins in diverse ways, to affect conversion of PrPC to its protease-resistant state. None affected conversion in the absence of pre-existing PrPSc. In its presence, only two, GroEL and Hsp104 (heat shock protein 104), significantly affected conversion. Both promoted it, but the reaction characteristics of conversions with the two chaperones were distinct. In contrast, chemical chaperones inhibited conversion. Our findings provide new mechanistic insights into nature of PrP conversions, and provide a new set of tools for studying the process underlying TSE pathogenesis.
Figures
Similar articles
-
Sulfated glycans and elevated temperature stimulate PrP(Sc)-dependent cell-free formation of protease-resistant prion protein.EMBO J. 2001 Feb 1;20(3):377-86. doi: 10.1093/emboj/20.3.377. EMBO J. 2001. PMID: 11157745 Free PMC article.
-
Cell-free formation of protease-resistant prion protein.Nature. 1994 Aug 11;370(6489):471-4. doi: 10.1038/370471a0. Nature. 1994. PMID: 7913989
-
Sheep scrapie susceptibility-linked polymorphisms do not modulate the initial binding of cellular to disease-associated prion protein prior to conversion.J Gen Virol. 2005 Sep;86(Pt 9):2627-2634. doi: 10.1099/vir.0.80901-0. J Gen Virol. 2005. PMID: 16099922
-
Prion protein conversion in vitro.J Mol Med (Berl). 2004 Jun;82(6):348-56. doi: 10.1007/s00109-004-0534-3. Epub 2004 Mar 10. J Mol Med (Berl). 2004. PMID: 15014886 Review.
-
[Unfolding chaperone as a prion protein relating molecule].Rinsho Shinkeigaku. 2003 Nov;43(11):817-9. Rinsho Shinkeigaku. 2003. PMID: 15152473 Review. Japanese.
Cited by
-
Formation of native prions from minimal components in vitro.Proc Natl Acad Sci U S A. 2007 Jun 5;104(23):9741-6. doi: 10.1073/pnas.0702662104. Epub 2007 May 29. Proc Natl Acad Sci U S A. 2007. PMID: 17535913 Free PMC article.
-
Chaperonins in disease: mechanisms, models, and treatments.Mol Pathol. 2002 Aug;55(4):209-13. doi: 10.1136/mp.55.4.209. Mol Pathol. 2002. PMID: 12147708 Free PMC article. Review.
-
[Transcription factor YY1 participates in activation transcription of the human ribosomal protein L11 gene].Mol Biol (Mosk). 2008 Jan-Feb;42(1):110-6. doi: 10.1134/s0026893308010160. Mol Biol (Mosk). 2008. PMID: 18389627 Russian.
-
Heparan Sulfate and Heparin Promote Faithful Prion Replication in Vitro by Binding to Normal and Abnormal Prion Proteins in Protein Misfolding Cyclic Amplification.J Biol Chem. 2016 Dec 16;291(51):26478-26486. doi: 10.1074/jbc.M116.745851. Epub 2016 Nov 7. J Biol Chem. 2016. PMID: 27821590 Free PMC article.
-
Methods for studying prion protein (PrP) metabolism and the formation of protease-resistant PrP in cell culture and cell-free systems. An update.Mol Biotechnol. 1999 Nov;13(1):45-55. doi: 10.1385/MB:13:1:45. Mol Biotechnol. 1999. PMID: 10934521
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
