

Probing the architecture of a simple kinetochore using DNA-protein crosslinking
- PMID: 9396745
- PMCID: PMC2132615
- DOI: 10.1083/jcb.139.6.1383
Probing the architecture of a simple kinetochore using DNA-protein crosslinking
Abstract
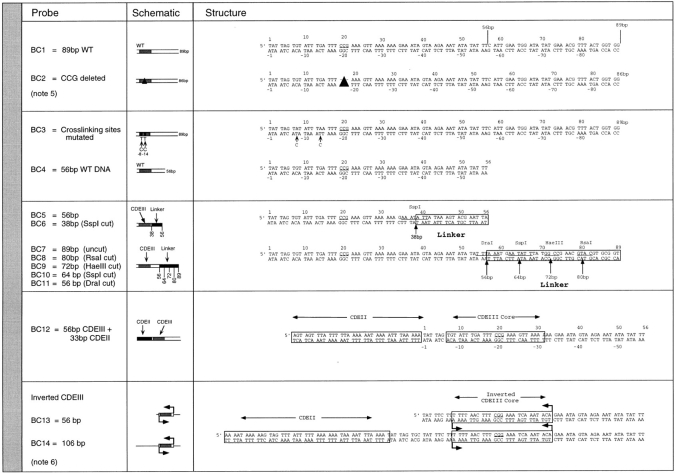
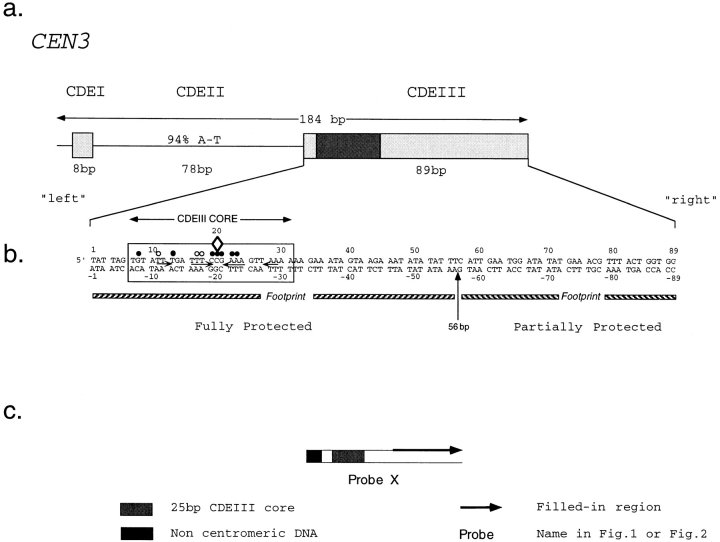
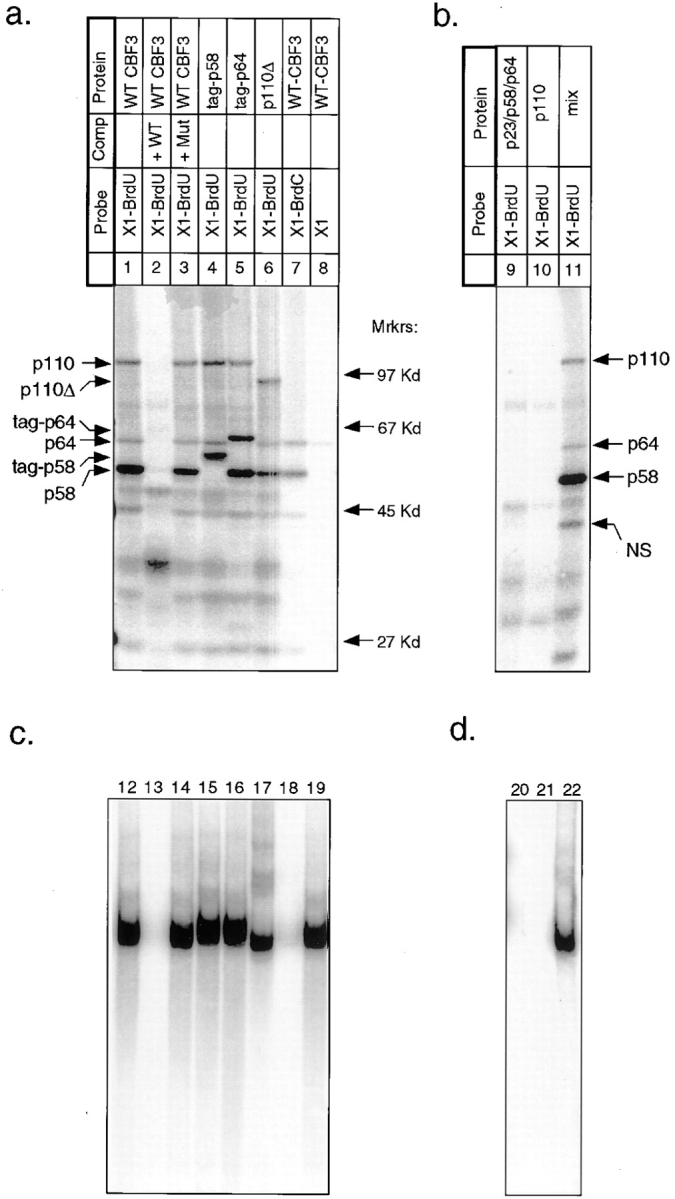
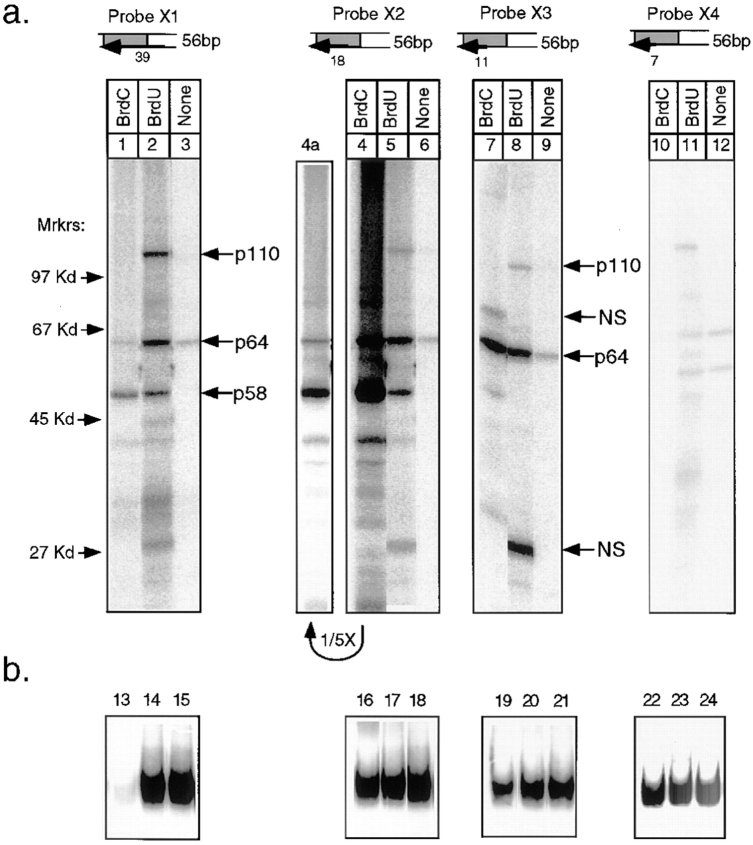
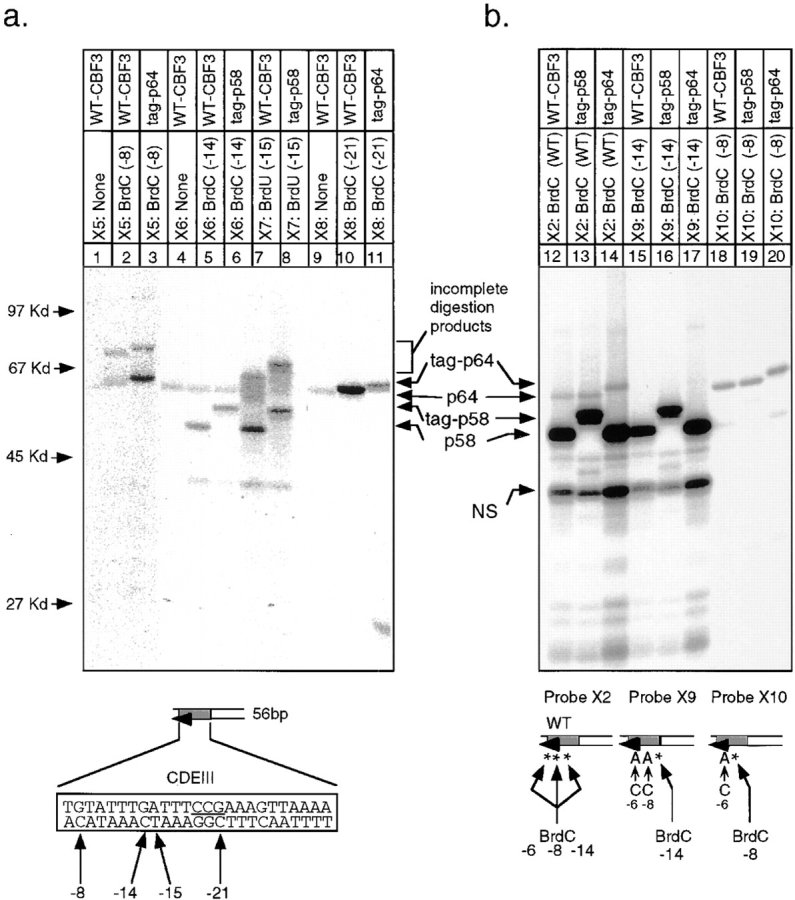
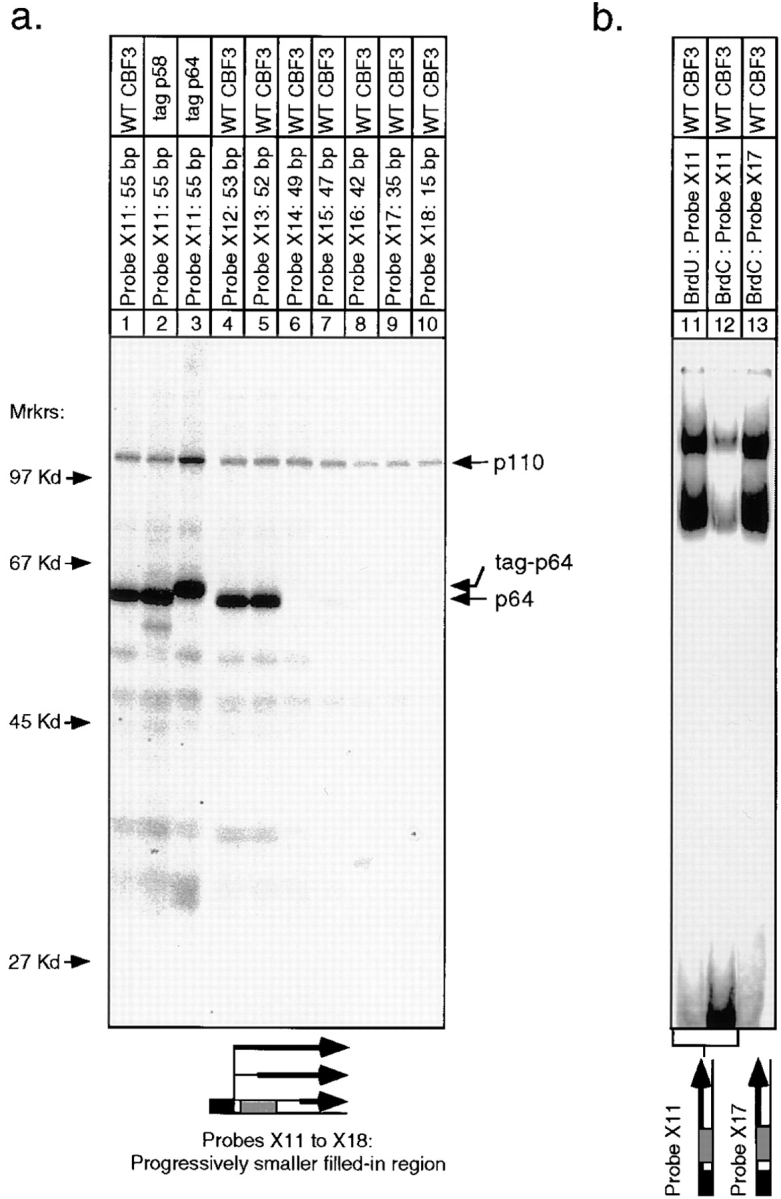
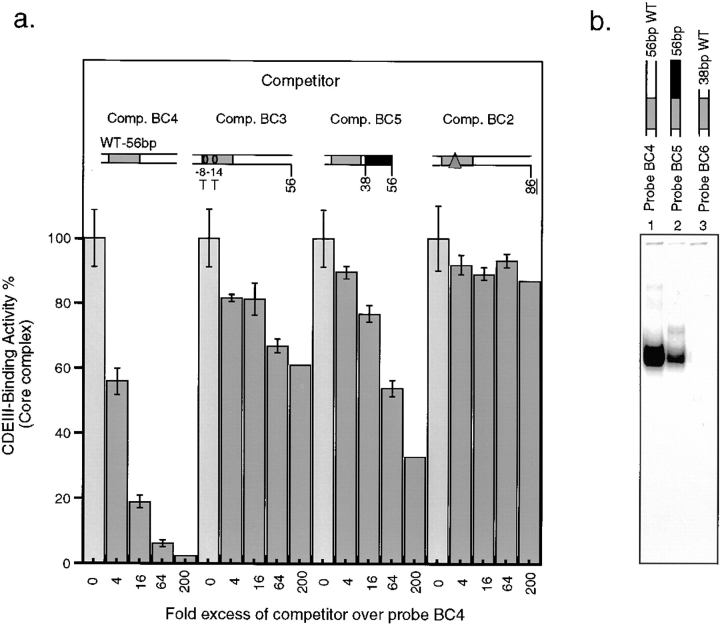
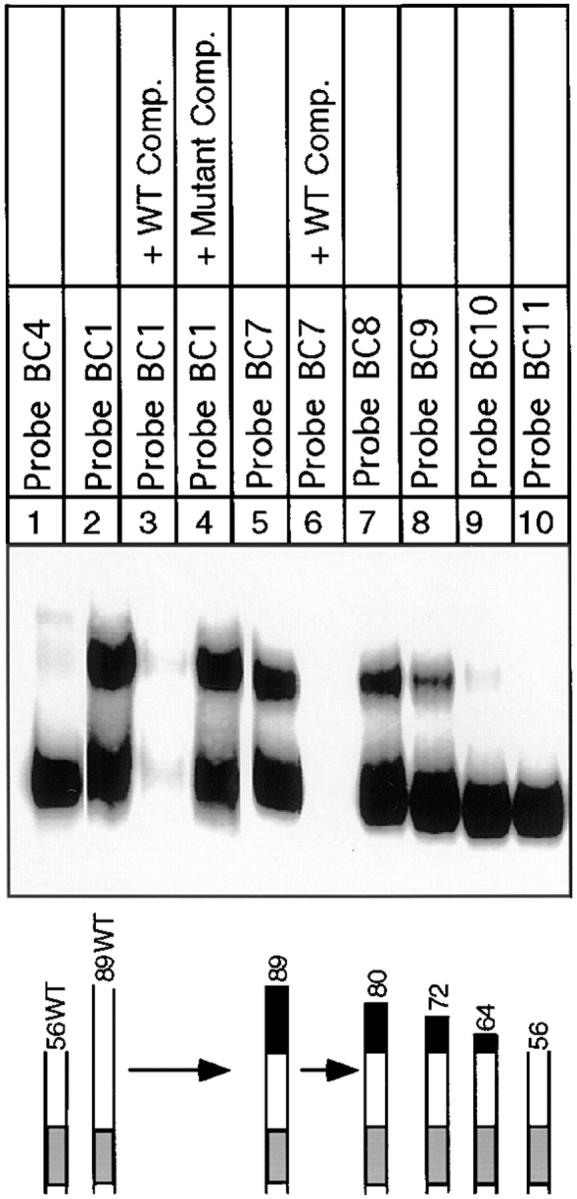
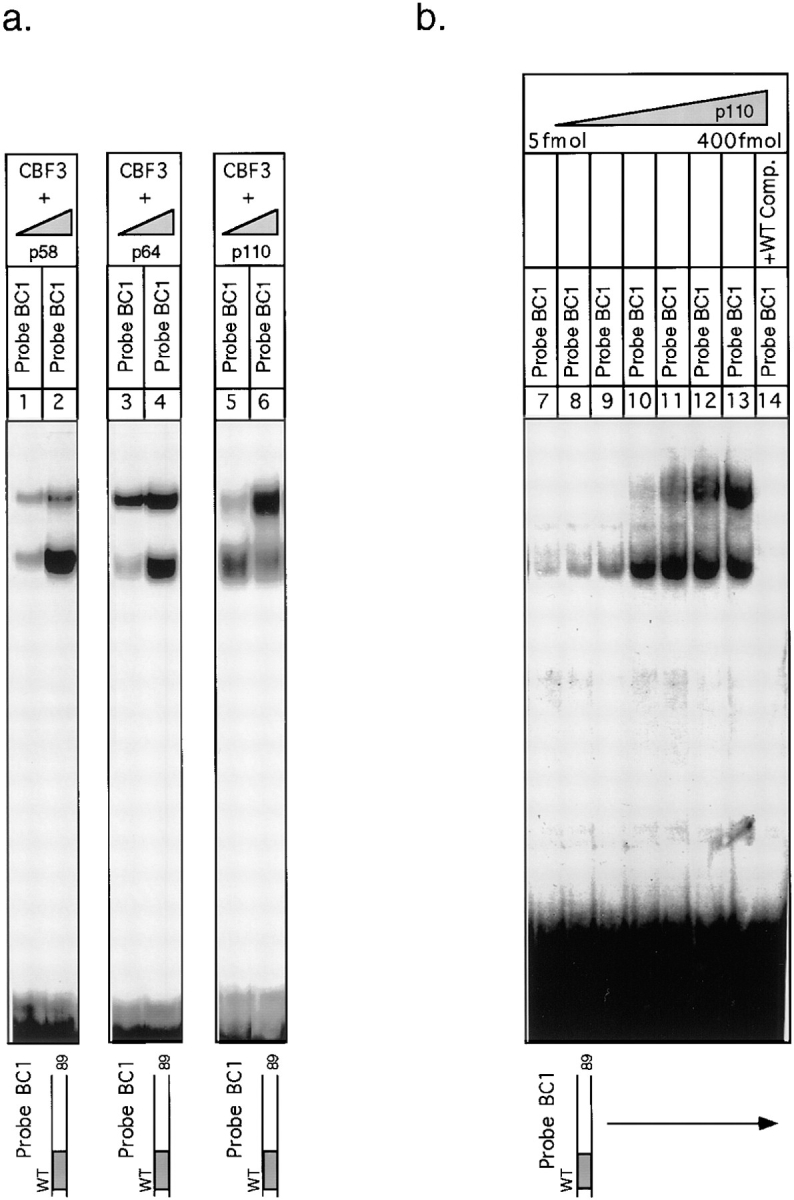
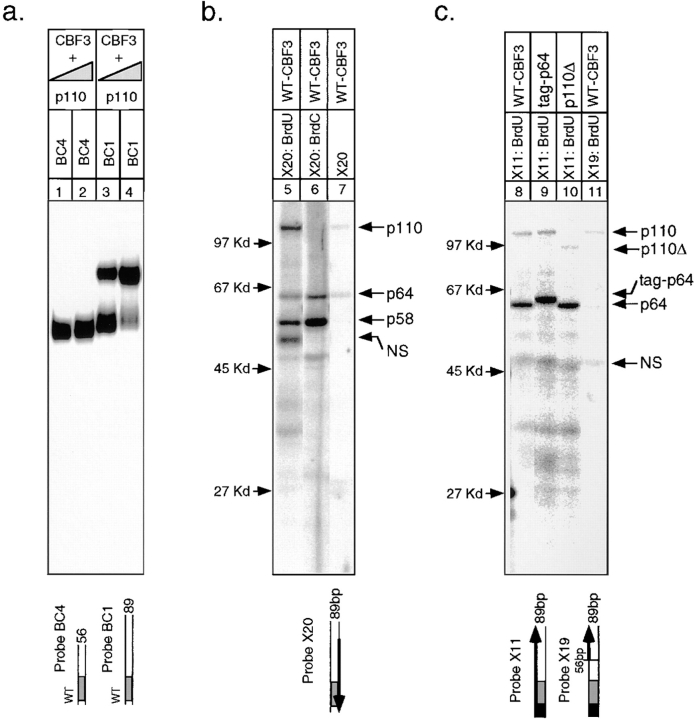
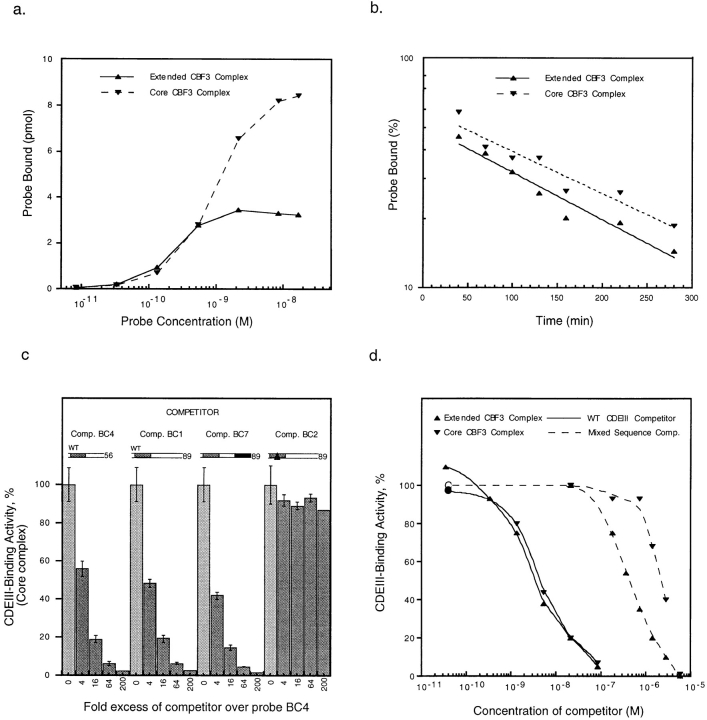
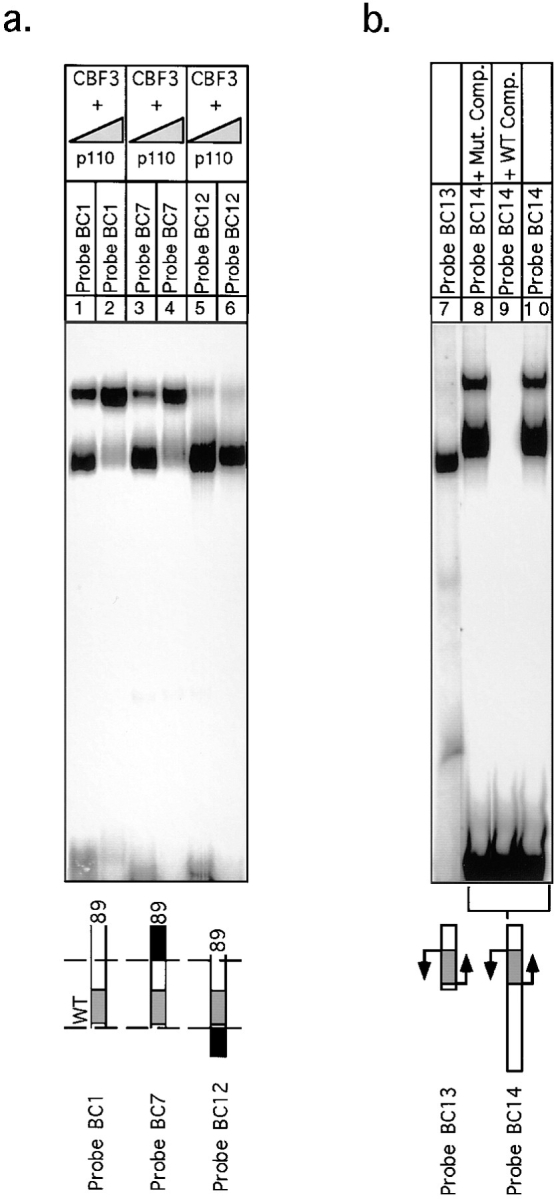
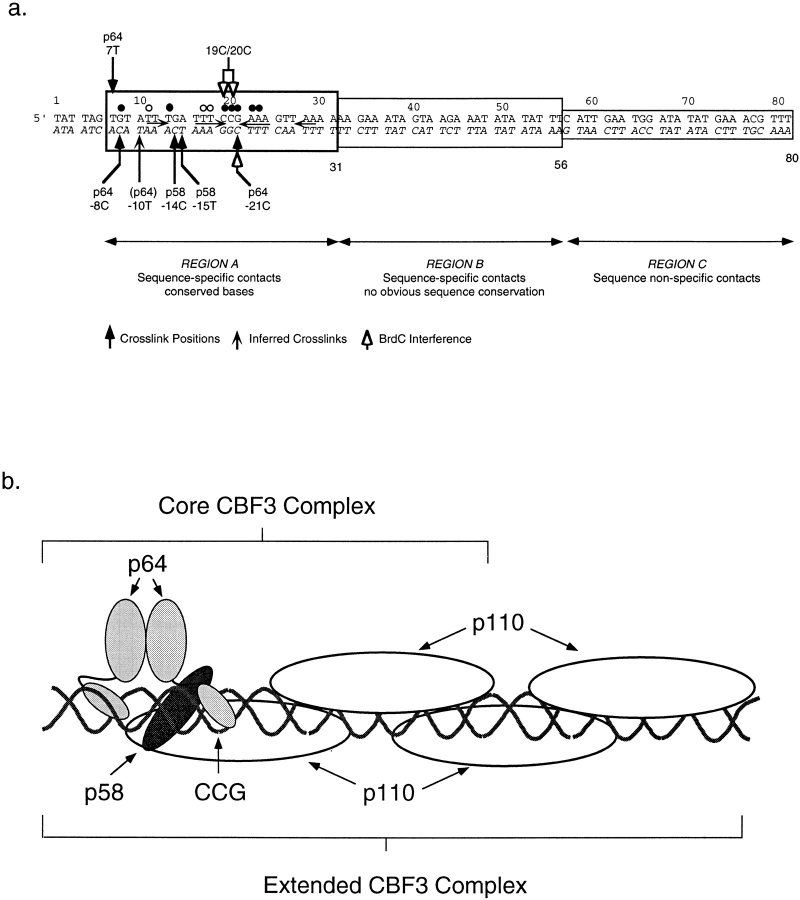
In budding yeast, accurate chromosome segregation requires that one and only one kinetochore assemble per chromosome. In this paper, we report the use of DNA-protein crosslinking and nondenaturing gel analysis to study the structure of CBF3, a four-protein complex that binds to the essential CDEIII region of Saccharomyces cerevisiae centromeres. We find that three subunits of CBF3 are in direct contact with CDEIII over a region of DNA that spans 80 bp. A highly asymmetric core complex containing p58(CTF13) p64(CEP3) and p110(NDC10) in direct contact with DNA forms at the genetically defined center of CDEIII. This core complex spans approximately 56 bp of CEN3. An extended complex comprising the core complex and additional DNA-bound p110(NDC10) also forms. It spans approximately 80 bp of DNA. CBF3 makes sequence-specific and -nonspecific contacts with DNA. Both contribute significantly to the energy of CBF3-DNA interaction. Moreover, important sequence-specific contacts are made with bases that are not conserved among yeast centromeres. These findings provide a foundation for understanding the organization of the CBF3-centromere complex, a structure that appears to initiate the formation of microtubule attachment sites at yeast kinetochores. These results also have implications for understanding centromere-binding proteins in higher cells.
Figures

References
-
- Bloom K, Carbon J. Yeast centromere DNA is in a unique and highly ordered structure in chromosomes and small circular minichromosomes. Cell. 1982;29:305–317. - PubMed
-
- Cai M, Davis RW. Yeast centromere binding protein CBF1, of the helix-loop-helix family, is required for chromosome stability and methionine prototrophy. Cell. 1990;61:437–446. - PubMed
-
- Clarke L. Centromeres of budding and fission yeast. TIG (Trends Genet) 1990;6:150–154. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
