

On the role of myosin-II in cytokinesis: division of Dictyostelium cells under adhesive and nonadhesive conditions
- PMID: 9398680
- PMCID: PMC25732
- DOI: 10.1091/mbc.8.12.2617
On the role of myosin-II in cytokinesis: division of Dictyostelium cells under adhesive and nonadhesive conditions
Abstract
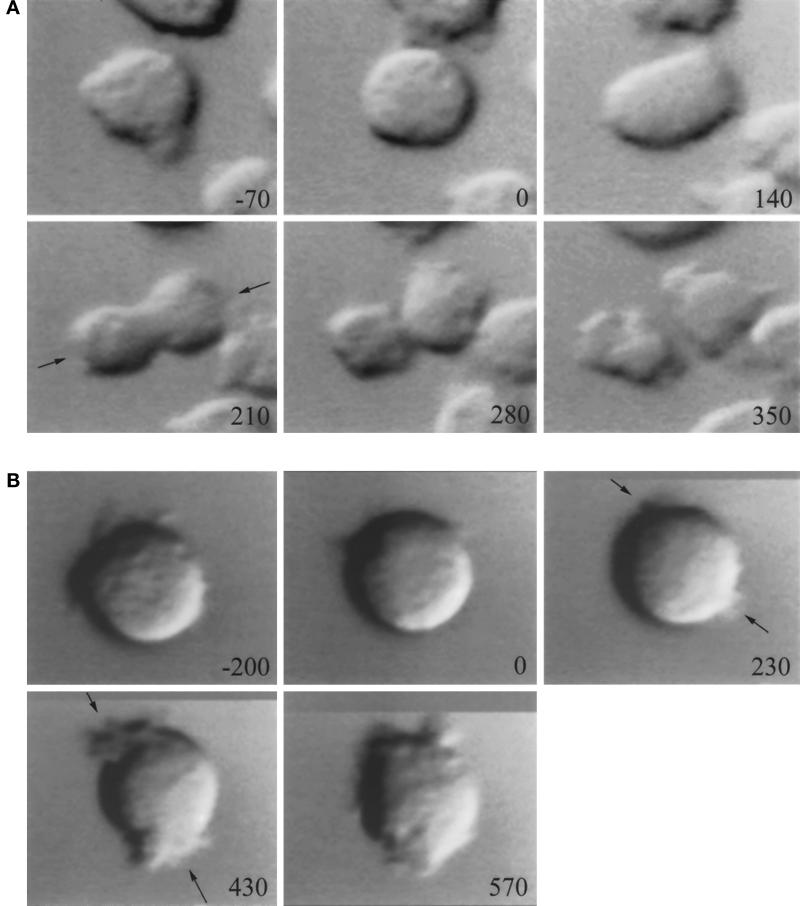
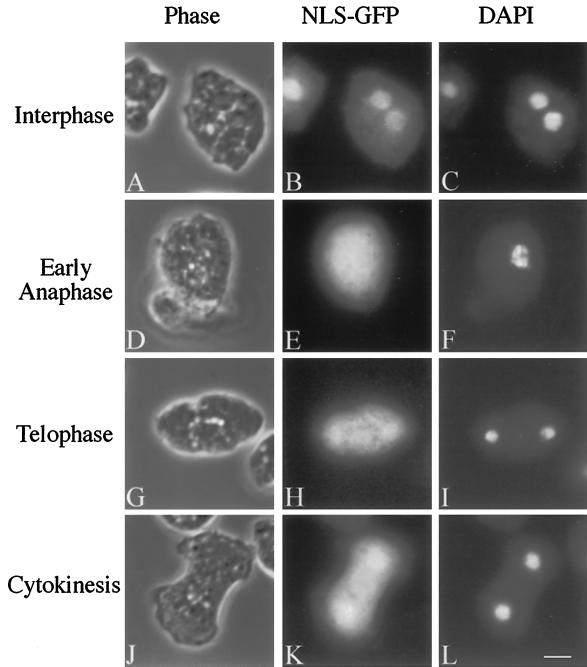
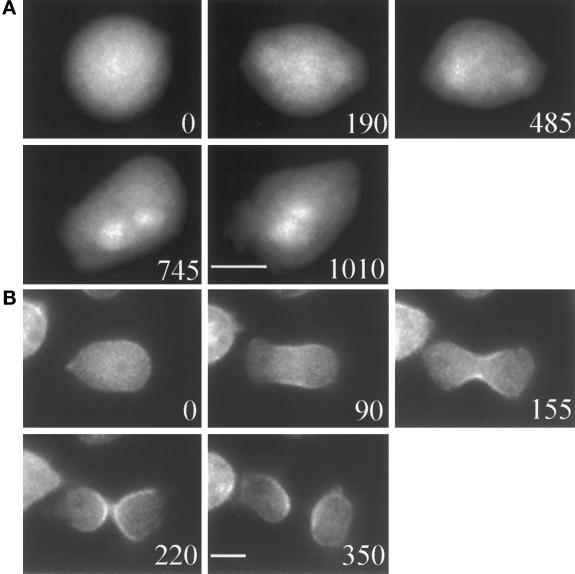
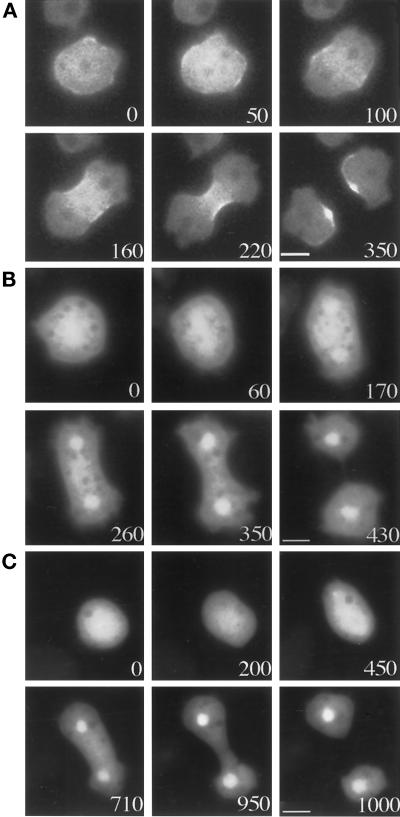
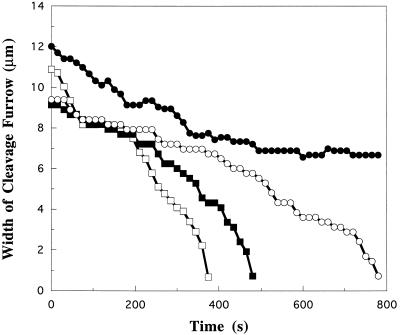
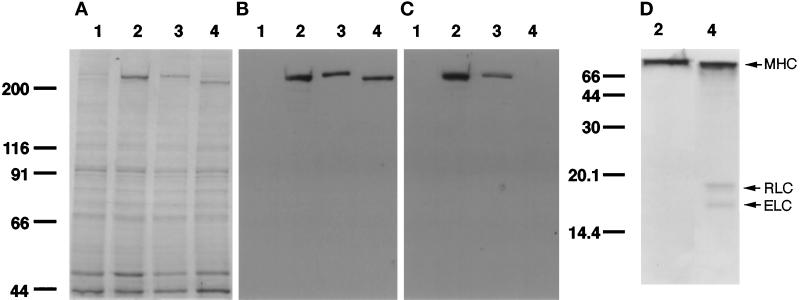
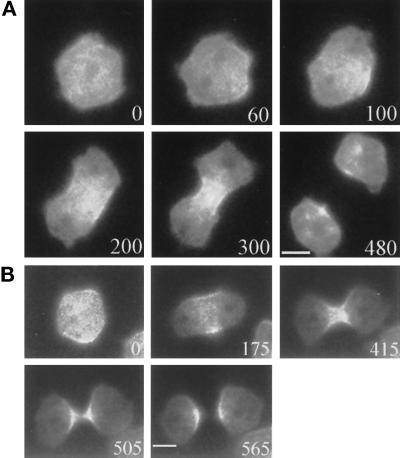
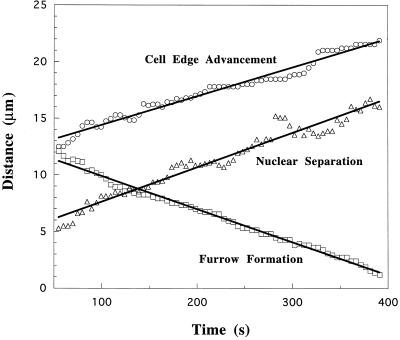
We have investigated the role of myosin in cytokinesis in Dictyostelium cells by examining cells under both adhesive and nonadhesive conditions. On an adhesive surface, both wild-type and myosin-null cells undergo the normal processes of mitotic rounding, cell elongation, polar ruffling, furrow ingression, and separation of daughter cells. When cells are denied adhesion through culturing in suspension or on a hydrophobic surface, wild-type cells undergo these same processes. However, cells lacking myosin round up and polar ruffle, but fail to elongate, furrow, or divide. These differences show that cell division can be driven by two mechanisms that we term Cytokinesis A, which requires myosin, and Cytokinesis B, which is cell adhesion dependent. We have used these approaches to examine cells expressing a myosin whose two light chain-binding sites were deleted (DeltaBLCBS-myosin). Although this myosin is a slower motor than wild-type myosin and has constitutively high activity due to the abolition of regulation by light-chain phosphorylation, cells expressing DeltaBLCBS-myosin were previously shown to divide in suspension (Uyeda et al., 1996). However, we suspected their behavior during cytokinesis to be different from wild-type cells given the large alteration in their myosin. Surprisingly, DeltaBLCBS-myosin undergoes relatively normal spatial and temporal changes in localization during mitosis. Furthermore, the rate of furrow progression in cells expressing a DeltaBLCBS-myosin is similar to that in wild-type cells.
Figures
References
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Burton K, Taylor DL. Traction forces of cytokinesis measured with optically modified elastic substrata. Nature. 1997;385:450–454. - PubMed
-
- De Lozanne A, Spudich JA. Disruption of the Dictyostelium myosin heavy chain gene by homologous recombination. Science. 1987;236:1086–1091. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
