

RGS2/G0S8 is a selective inhibitor of Gqalpha function
- PMID: 9405622
- PMCID: PMC24991
- DOI: 10.1073/pnas.94.26.14389
RGS2/G0S8 is a selective inhibitor of Gqalpha function
Abstract
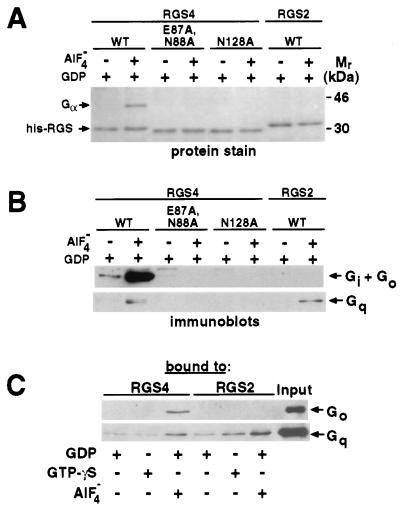
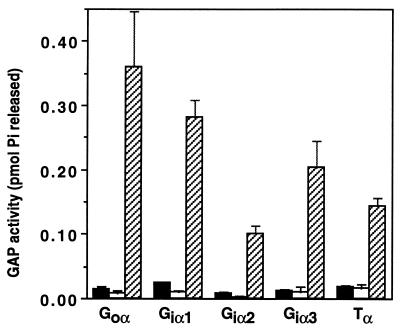
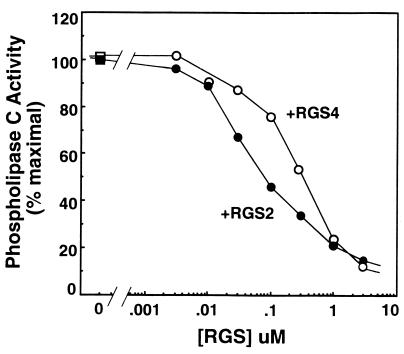
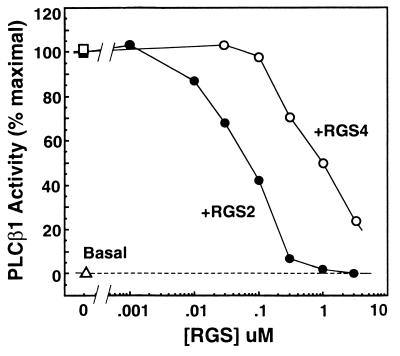
RGS (regulators of G protein signaling) proteins are GTPase activating proteins that inhibit signaling by heterotrimeric G proteins. All RGS proteins studied to date act on members of the Gialpha family, but not Gsalpha or G12alpha. RGS4 regulates Gialpha family members and Gqalpha. RGS2 (G0S8) is exceptional because the G proteins it regulates have not been identified. We report that RGS2 is a selective and potent inhibitor of Gqalpha function. RGS2 selectively binds Gqalpha, but not other Galpha proteins (Gi, Go, Gs, G12/13) in brain membranes; RGS4 binds Gqalpha and Gialpha family members. RGS2 binds purified recombinant Gqalpha, but not Goalpha, whereas RGS4 binds either. RGS2 does not stimulate the GTPase activities of Gsalpha or Gialpha family members, even at a protein concentration 3000-fold higher than is sufficient to observe effects of RGS4 on Gialpha family members. In contrast, RGS2 and RGS4 completely inhibit Gq-directed activation of phospholipase C in cell membranes. When reconstituted with phospholipid vesicles, RGS2 is 10-fold more potent than RGS4 in blocking Gqalpha-directed activation of phospholipase Cbeta1. These results identify a clear physiological role for RGS2, and describe the first example of an RGS protein that is a selective inhibitor of Gqalpha function.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
