

Benthic-pelagic links and rocky intertidal communities: bottom-up effects on top-down control?
- PMID: 9405647
- PMCID: PMC25044
- DOI: 10.1073/pnas.94.26.14530
Benthic-pelagic links and rocky intertidal communities: bottom-up effects on top-down control?
Abstract
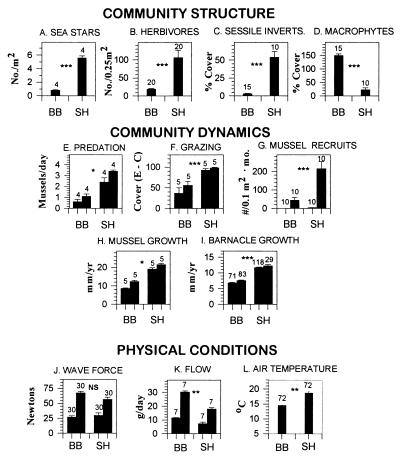
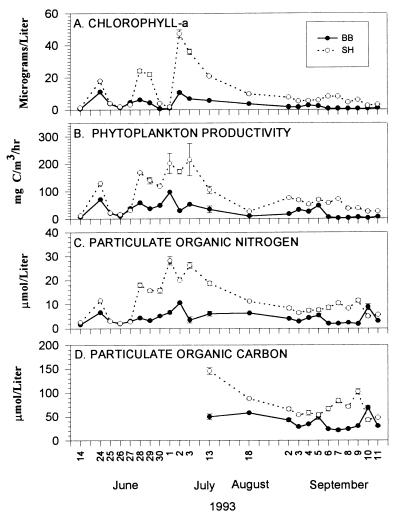
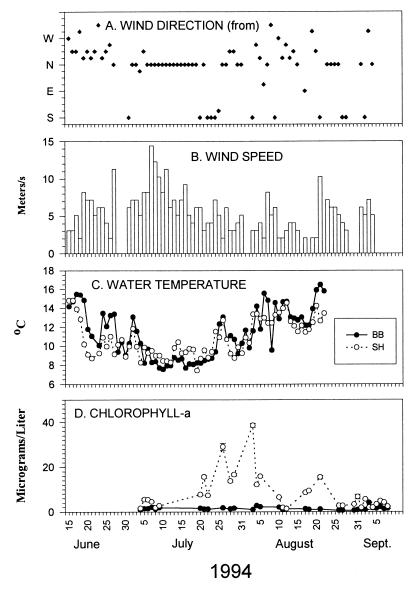
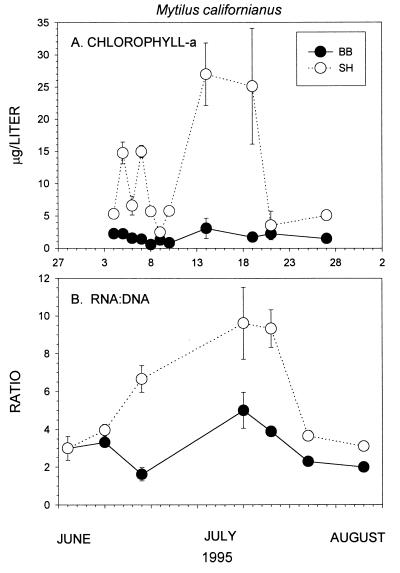
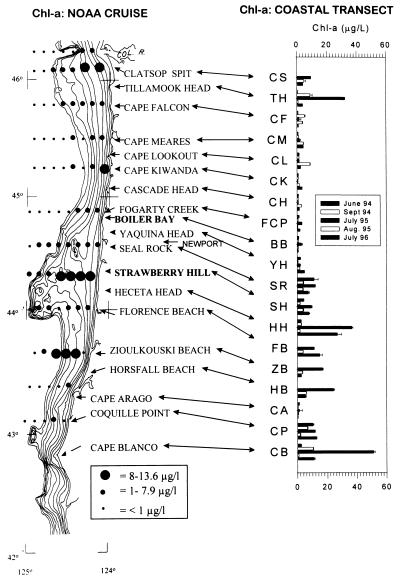
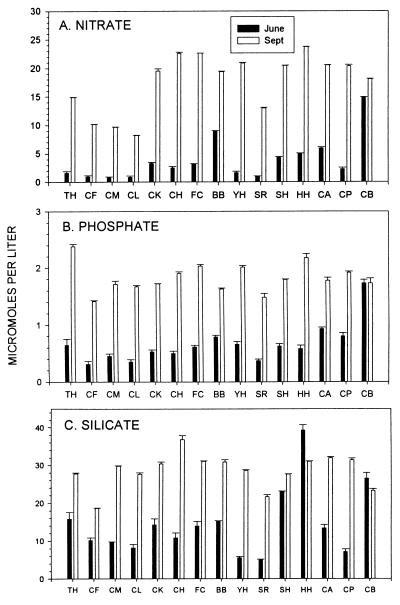
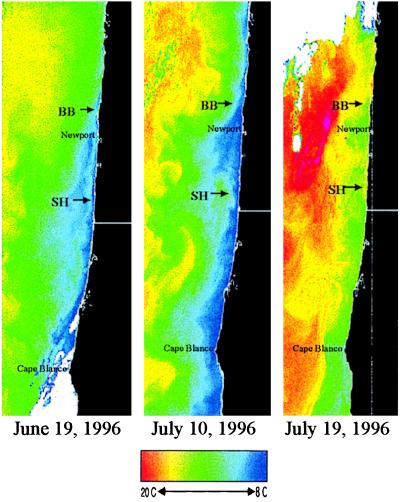
Insight into the dependence of benthic communities on biological and physical processes in nearshore pelagic environments, long considered a "black box," has eluded ecologists. In rocky intertidal communities at Oregon coastal sites 80 km apart, differences in abundance of sessile invertebrates, herbivores, carnivores, and macrophytes in the low zone were not readily explained by local scale differences in hydrodynamic or physical conditions (wave forces, surge flow, or air temperature during low tide). Field experiments employing predator and herbivore manipulations and prey transplants suggested top-down (predation, grazing) processes varied positively with bottom-up processes (growth of filter-feeders, prey recruitment), but the basis for these differences was unknown. Shore-based sampling revealed that between-site differences were associated with nearshore oceanographic conditions, including phytoplankton concentration and productivity, particulates, and water temperature during upwelling. Further, samples taken at 19 sites along 380 km of coastline suggested that the differences documented between two sites reflect broader scale gradients of phytoplankton concentration. Among several alternative explanations, a coastal hydrodynamics hypothesis, reflecting mesoscale (tens to hundreds of kilometers) variation in the interaction between offshore currents and winds and continental shelf bathymetry, was inferred to be the primary underlying cause. Satellite imagery and offshore chlorophyll-a samples are consistent with the postulated mechanism. Our results suggest that benthic community dynamics can be coupled to pelagic ecosystems by both trophic and transport linkages.
Figures
References
-
- Levin S A. Ecology. 1992;73:1943–1967.
-
- Persson L, Bengtsson J, Menge B A, Power M E. In: Food Webs: Integration of Patterns & Dynamics. Polis G A, Winemiller K O, editors. New York: Chapman and Hall; 1995. pp. 396–434.
-
- Carpenter S R, editor. Complex Interactions in Lake Communities. New York: Springer; 1988.
-
- Paine R T, Levin S A. Ecol Monogr. 1981;51:145–178.
-
- Duggins D O, Simenstad C A, Estes J A. Science. 1989;245:170–173. - PubMed
LinkOut - more resources
Full Text Sources
