

A new mechanism of sound generation in songbirds
- PMID: 9405691
- PMCID: PMC25115
- DOI: 10.1073/pnas.94.26.14787
A new mechanism of sound generation in songbirds
Abstract
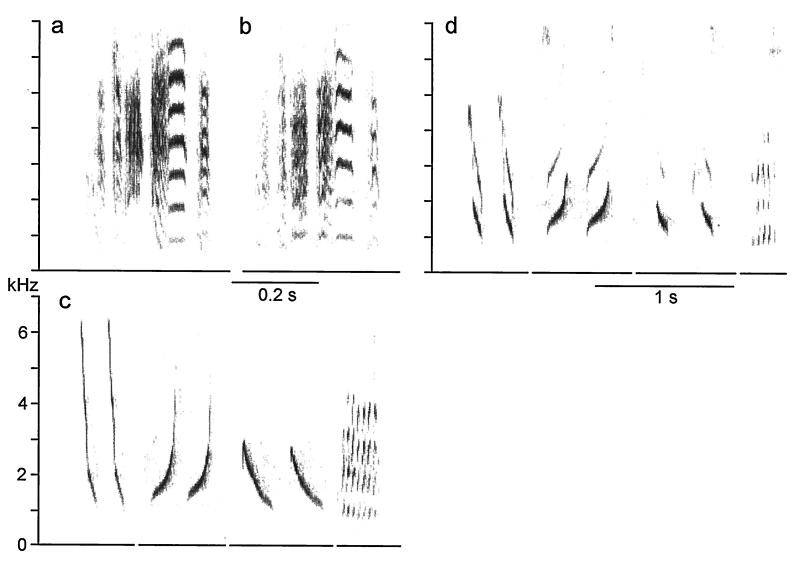
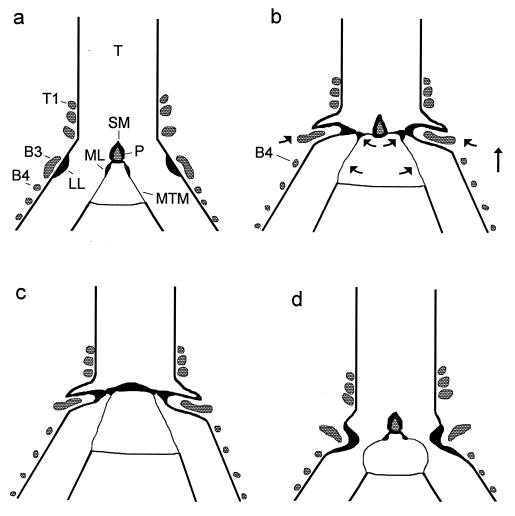
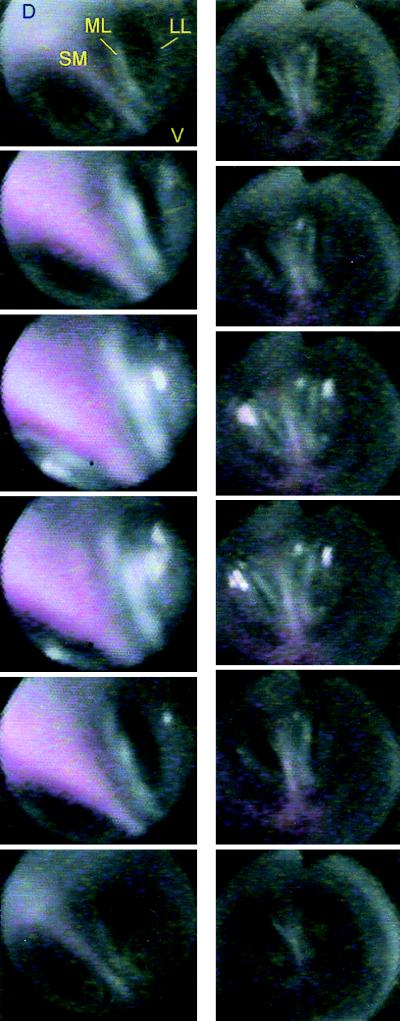
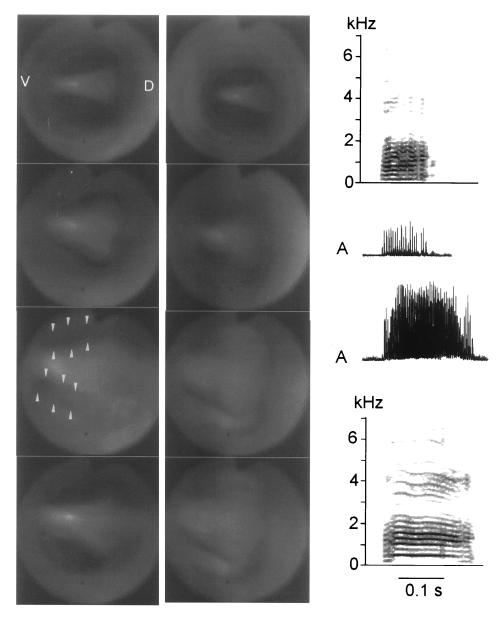
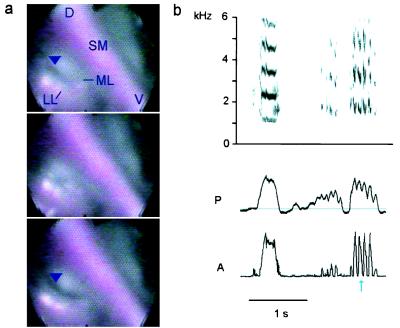
Our current understanding of the sound-generating mechanism in the songbird vocal organ, the syrinx, is based on indirect evidence and theoretical treatments. The classical avian model of sound production postulates that the medial tympaniform membranes (MTM) are the principal sound generators. We tested the role of the MTM in sound generation and studied the songbird syrinx more directly by filming it endoscopically. After we surgically incapacitated the MTM as a vibratory source, zebra finches and cardinals were not only able to vocalize, but sang nearly normal song. This result shows clearly that the MTM are not the principal sound source. The endoscopic images of the intact songbird syrinx during spontaneous and brain stimulation-induced vocalizations illustrate the dynamics of syringeal reconfiguration before phonation and suggest a different model for sound production. Phonation is initiated by rostrad movement and stretching of the syrinx. At the same time, the syrinx is closed through movement of two soft tissue masses, the medial and lateral labia, into the bronchial lumen. Sound production always is accompanied by vibratory motions of both labia, indicating that these vibrations may be the sound source. However, because of the low temporal resolution of the imaging system, the frequency and phase of labial vibrations could not be assessed in relation to that of the generated sound. Nevertheless, in contrast to the previous model, these observations show that both labia contribute to aperture control and strongly suggest that they play an important role as principal sound generators.
Figures
References
-
- Konishi M. Annu Rev Neurosci. 1985;8:125–170. - PubMed
-
- Nottebohm F. J Comp Physiol A. 1996;179:149–156. - PubMed
-
- Marler P. Am Scientist. 1970;58:669–673. - PubMed
-
- Hauser M D. The Evolution of Communication. Cambridge, MA: MIT; 1996.
-
- Fink B R. The Human Larynx: A Functional Study. New York: Raven; 1975.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
