

doi: 10.1101/gad.11.24.3306.
5'-Capping enzymes are targeted to pre-mRNA by binding to the phosphorylated carboxy-terminal domain of RNA polymerase II
Affiliations
- PMID: 9407024
- PMCID: PMC316822
- DOI: 10.1101/gad.11.24.3306
Item in Clipboard
5'-Capping enzymes are targeted to pre-mRNA by binding to the phosphorylated carboxy-terminal domain of RNA polymerase II
Genes Dev.
.
Abstract
We have investigated the role of the RNA Polymerase II (Pol II) carboxy-terminal domain (CTD) in mRNA 5' capping. Transcripts made in vivo by Pol II with a truncated CTD had a lower proportion of capped 5' ends than those made by Pol II with a full-length CTD. In addition, the enzymes responsible for cap synthesis, RNA guanylyltransferase, and RNA (guanine-7)-methyltransferase bound directly to the phosphorylated, but not to the nonphosphorylated, form of the CTD in vitro. These results suggest that: (1) Pol II-specific capping of nascent transcripts in vivo is enhanced by recruitment of the capping enzymes to the CTD and (2) capping is co-ordinated with CTD phosphorylation.
Figures
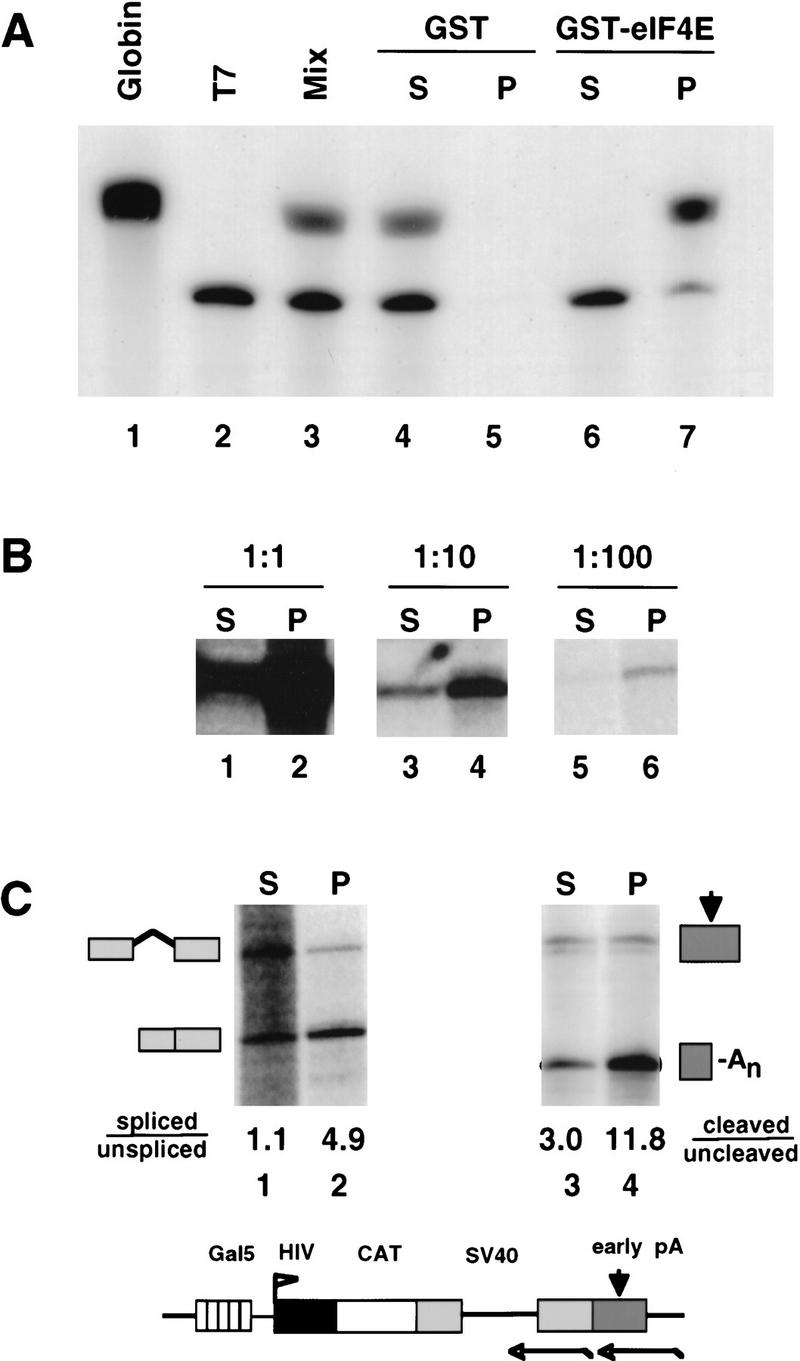
GST–eIF4E pull-down assay of capped RNAs. (A) A mixture (lane 3) of 3H-labeled rabbit globin mRNA (GIBCO-BRL) (capped, lane 1) and a synthetic T7 transcript (uncapped, lane 2) were fractionated using GST (lanes 4,5) or GST–eIF4E (lanes 6,7). Supernatant (S) and pellet (P) fractions were analyzed by electrophoresis and fluorography. (B) RNA from α-amanitin-treated 293 cells transfected with HA–WT α-amanitin-resistant Pol II, pGal5 HIV2 CAT reporter, and Gal4–VP16 activator was serially diluted with RNA from untransfected cells, then fractionated by GST–eIF4E pull-down. A total of 5 μg of RNA was used for each pull-down. Supernatant (S) and pellet (P) fractions were analyzed by RNase protection with a probe spanning the start site (Fig. 2C). The 160-base protection product corresponding to correct HIV2 5′ ends is shown. (C) RNA in the GST–eIF4E supernatant (S) fraction is spliced and polyadenylated less well than RNA in the pellet (P) fraction. GST–eIF4E fractionated RNA from α-amanitin-treated 293 cells expressing HA–WT Pol II and pGal5 HIV2 CAT activated by Gal4–VP16 was analyzed for splicing of the SV40 t intron (lanes 1,2) and cleavage at the SV40 early poly(A) site (lanes 3,4) using antisense RNA probes as indicated in the map. Processed/unprocessed ratios are given after correction for the 32P-labeled U content of the RNase protection products. A map and RNase protection strategy for the pGal5 HIV2 CAT reporter gene is shown.
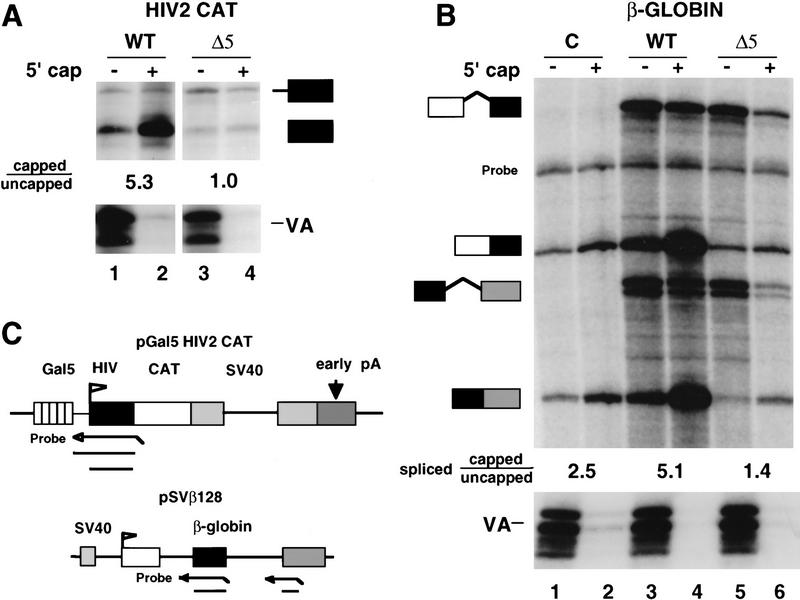
CTD truncation reduces the ratio of capped/uncapped 5′ ends. (A) Analysis of WT and Δ5 transcripts of pGal5 HIV2 CAT by RNase protection as diagrammed. RNA (20 μg) from α-amanitin-treated cells was separated into capped (+) and uncapped (−) fractions and analyzed by RNase protection with probes for the the HIV2 5′ end and VA1. Capped/uncapped ratios for correctly initiated RNAs (lower band) are marked. (B) Analysis of human β-globin RNA transcribed by wild-type (WT) and CTD-truncated (Δ5) α-amanitin-resistant Pol II. The pSVβ128 reporter and Pol II expression vectors or CMV–neo control (C) were transfected transiently into 293 cells. GST–eIF4E fractionated RNA (20 μg) was analyzed as in A by RNase protection with probes for VA control, β-globin intron 1, and intron 2 as diagrammed in C. A band corresponding to incompletely digested intron 2 probe is marked. The capped/uncapped ratios shown are average values for the two “spliced” RNase protected bands. The spliced/unspliced ratios for intron 1 after correcting for 32P-labeled U content are 1.6, 11.5, 0.4, and 2.1 for lanes 3–6, respectively. The spliced/unspliced ratios for intron 2 are 0.7, 5.8, 0.2, and 1.2 for lanes 3–6. (C) Maps and RNase protection strategies for the pGal5 HIV2 CAT and pSVβ128 β-globin reporter genes.
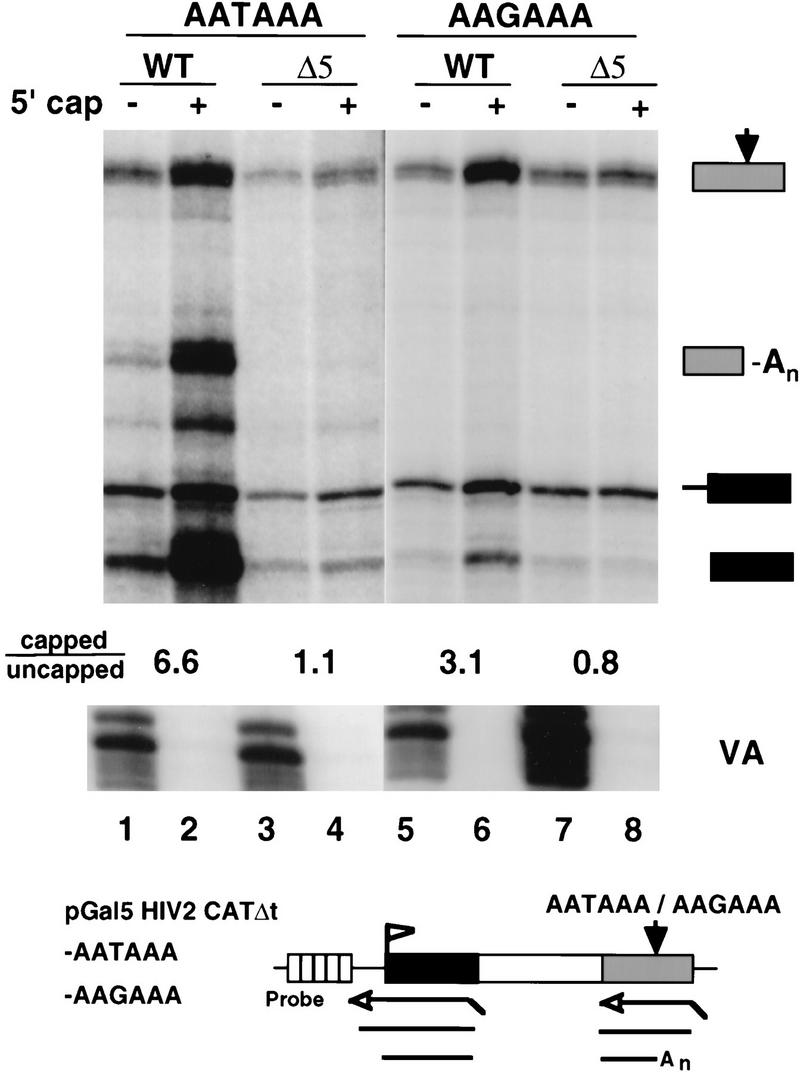
CTD truncation reduces capping of unspliced and unpolyadenylated RNAs. RNase protection of intronless CAT transcripts from pGal5 HIV2 CATΔt–AATAAA and pGal5 HIV2 CATΔt–AAGAAA transcribed by WT or Δ5 Pol II. Total RNA (6 μg) from transfected 293 cells was fractionated by GST–eIF4E binding and analyzed with 5′ and 3′ antisense probes shown in the diagram. The capped/uncapped ratios are shown for correctly initiated transcripts (marked with a solid box). The VA control is indicated. At the poly(A) site, the ratios of cleaved/uncleaved transcripts after correcting for 32P-labeled U content of the RNase protection products are 0.7, 2.1, <0.3, and <0.3 for lanes 1–4, respectively.
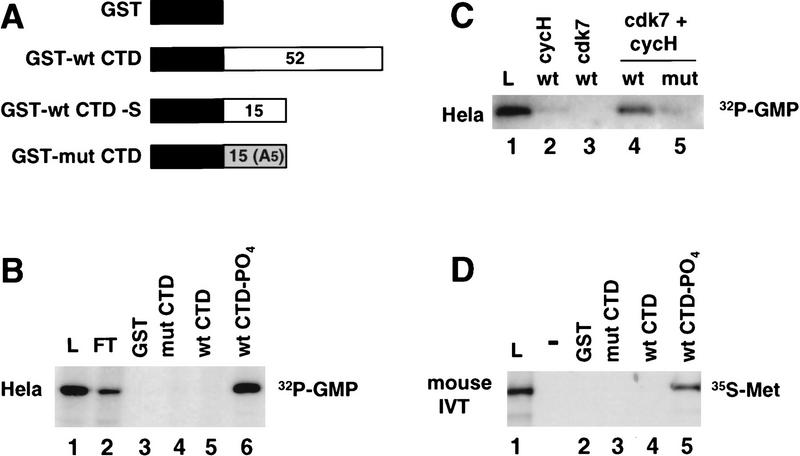
Mammalian guanylyltransferase binds specifically to the phosphorylated CTD. (A) GST–CTD fusion proteins used for affinity chromatography. The number of heptad repeats in each is shown. (B) HeLa guanylyltransferase binds to the phosphorylated CTD. Nuclear extract was chromatographed sequentially on GST, GST–mut CTD, GST–wt CTD, and GST–wt CTD–PO4 that had been phosphorylated in HeLa nuclear extract and extensively washed in high-salt buffer (see Materials and Methods). GST–mut CTD was used at a higher ligand concentration to compensate for its shorter length. Load (L), flowthrough (FT), and high-salt eluates were assayed for the 68-kD guanylyltransferase by [32P]GMP labeling. 0.033% of L and FT fractions were analyzed and 0.4% of the eluates. (C) GST–wt CTD and GST–mut CTD were phosphorylated in reactions with baculoviral cyclin H, cdk7, or both cdk7 and cycH. HeLa extract was bound to these resins and the load (2%) and high-salt eluates (5%) as assayed for guanylyltransferase. (D) Specific binding of in vitro-translated mouse guanylyltransferase to the phosphorylated CTD. [35S]Met-labeled in vitro translation product of amino acids 1–590 of mouse guanylyltransferase homolog (see Fig. 5) was bound to GST–CTD resins. Load (L; 5% of total) and high-salt eluates (30%) from the resins were electrophoresed on an SDS gel and autoradiographed.
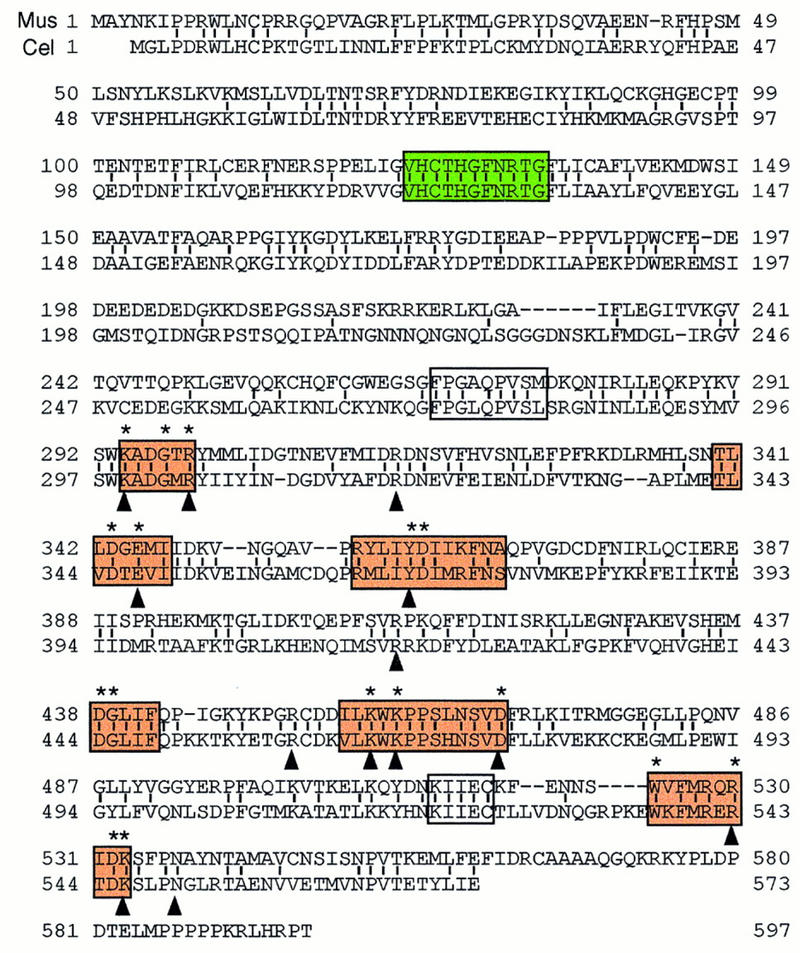
Sequence alignment of the mouse and C. elegans capping enzymes. The predicted mouse sequence was derived from two overlapping cDNA clones. The C. elegans sequence is from Takagi et al. (1997) and Wang et al. (1997). Amino acid identity, denoted by vertical lines, is 43%. Amino acids defined by alanine-scanning mutagenesis as essential for the activity of the S. cerevisiae capping enzyme Ceg1 are denoted by asterisks above the aligned sequences. The six nucleotidyl transferase superfamily motifs (I, III, IIIa, IV, V, and VI) are shown in bold black boxes. Motifs P (FPG × QPVS ×) and Vc (KIIEC) are in fine black boxes. The phosphatase motif (VHCTHGFNRTG) is in a gray box. Residues in proximity to the GTP moiety in the Chlorella virus capping enzyme–GTP cocrystal are indicated by arrowheads (Hakansson et al. 1997).
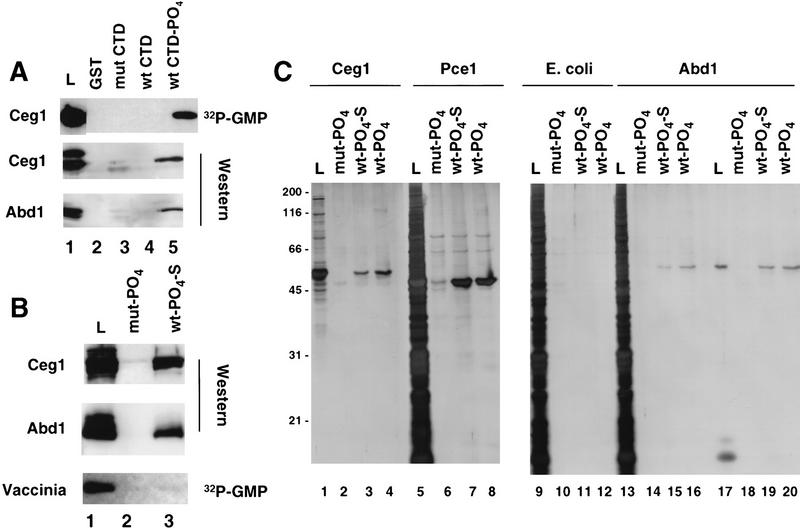
Yeast capping enzymes bind directly to the phosphorylated CTD. (A) Yeast guanylyltransferase, Ceg-1, methyltransferase, and Abd-1 bind to the phosphorylated CTD. Yeast extract partially purified on BioRex-70 was incubated with the GST–CTD resins, and the load (0.5%) and high-salt eluates (2%) were assayed by [32P]GMP labeling and Western blotting with antisera against Ceg1 and Abd1. Fivefold more protein was used for the Western blots. Several nonspecific bands detected by the antibodies in the load fraction were not bound to the CTD resins. (B) GST–CTD mutant (mut, lane 2) and wild-type (wt; 15 heptad repeats, lane 3) were phosphorylated with HeLa extract, washed extensively (see Materials and Methods), and used as ligands for affinity chromatography with bacterial fractions containing recombinant Ceg1, Abd1, or purified vaccinia capping enzyme. In lanes 1–6 the load (L, 0.74% of total, lane 1) and high salteluates (1.34%, lanes 2,3) were analyzed by Western blotting with anti-Ceg1 and Abd1 antisera. Vaccinia virus capping enzyme (4.7 units loaded, BRL) was assayed by [α32P]GTP labeling of the load (6% of total) and high-salt eluates (5%). (C) Silver-stained SDS gels of bacterially expressed yeast capping enzymes chromatographed on phosphorylated GST–CTD resins. Load (L; 10% of total) and high-salt eluates (20%) of recombinant S. cerevisiae and S. pombe guanylyltransferases (Ceg1 and Pce1) and S. cerevisiae methyltransferase (Abd1) were analyzed. As a negative control, total E. coli extract from a nonexpressing strain was used (lanes 9–12). Abd1 was chromatographed as a highly purified fraction (lanes 17–20) or after mixing with E. coli extract (lanes 13–16).
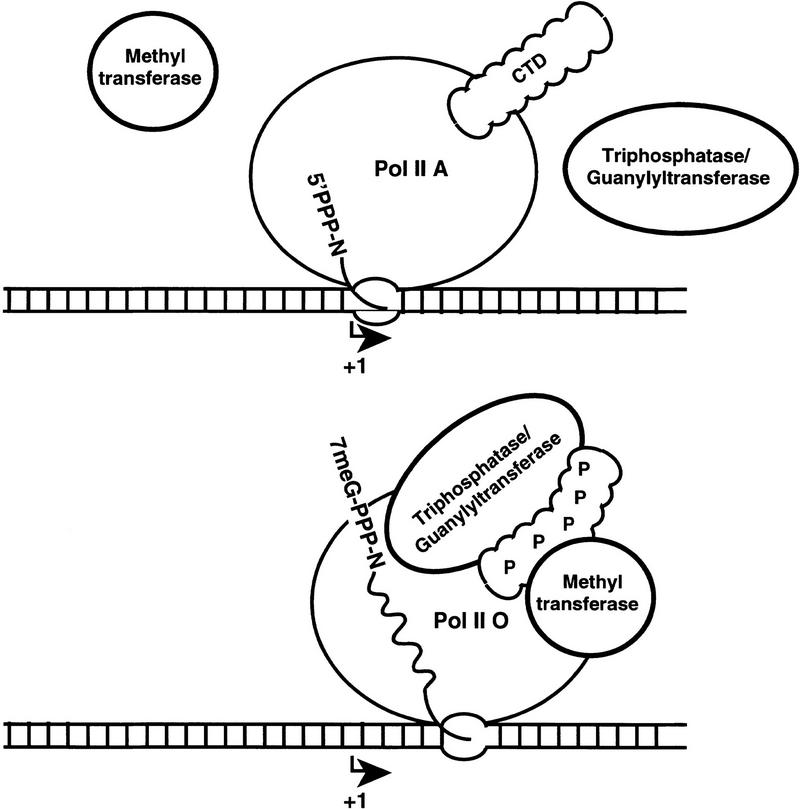
Model for recruitment of capping enzymes to the phosphorylated CTD.
Comment in
-
Transcription units as RNA processing units.Genes Dev. 1997 Dec 15;11(24):3279-85. doi: 10.1101/gad.11.24.3279. Genes Dev. 1997. PMID: 9407022 Review. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous