

DNA damage induces phosphorylation of the amino terminus of p53
- PMID: 9407038
- PMCID: PMC316806
- DOI: 10.1101/gad.11.24.3471
DNA damage induces phosphorylation of the amino terminus of p53
Abstract
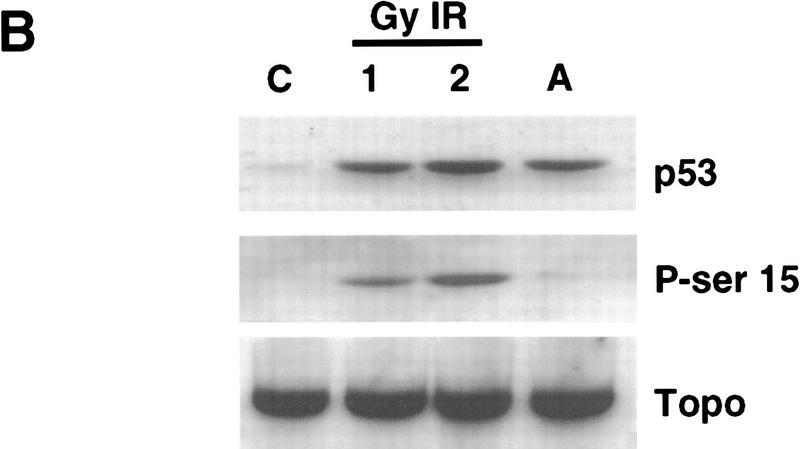
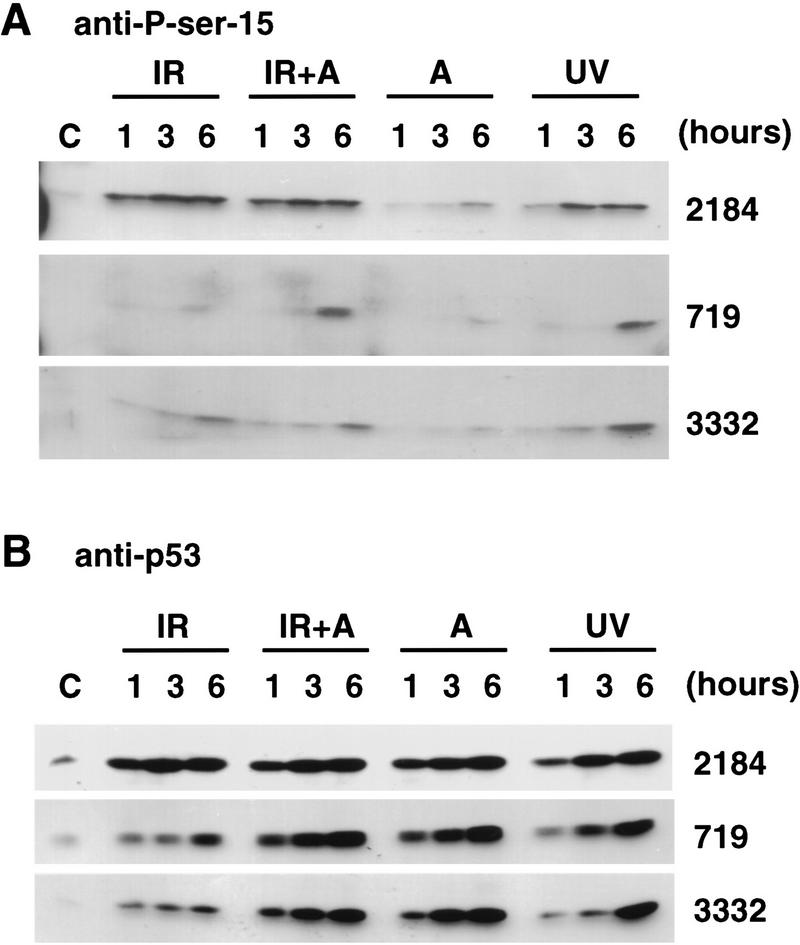
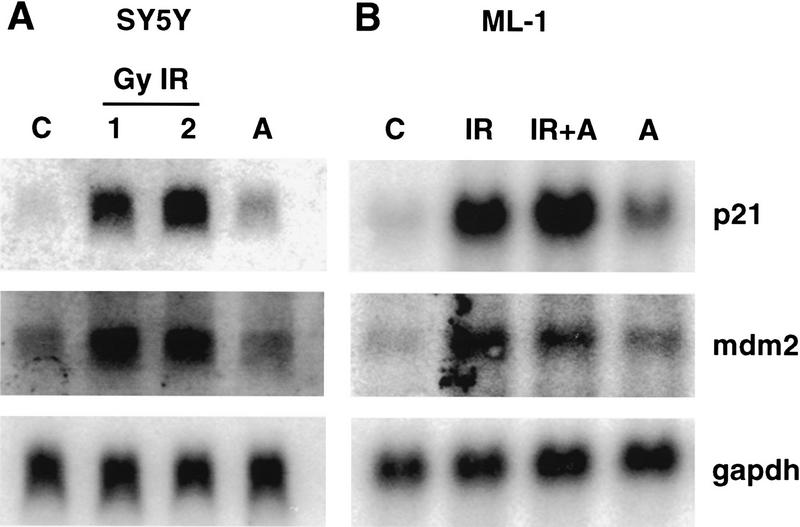
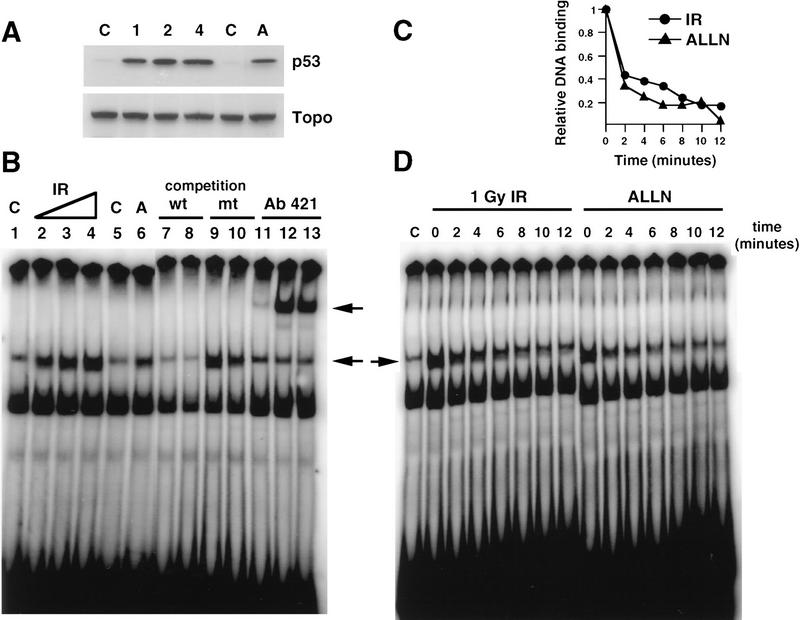
Data are presented demonstrating that DNA damage leads to specific post-translational modifications of p53 protein. Using two-dimensional peptide mapping of in vivo radiolabeled p53 tryptic phosphopeptides, recombinant truncated p53 protein, and synthetic p53 tryptic peptides, a unique p53 phosphopeptide was identified after exposure of ML-1 cells to ionizing irradiation. This peptide represents the first 24 amino acids of p53 and contains three phosphorylated serine residues. A specific p53 phosphopeptide antibody identified serine-15 as one of the two serines in p53 that becomes phosphorylated following DNA damage induced by either ionizing irradiation (IR) or ultraviolet (UV) irradiation in multiple cell types. IR-induced phosphorylation of p53 does not affect the kinetics of p53 binding to or dissociating from DNA as assessed by electrophoretic mobility-shift assays. However, p53 phosphorylation induced by DNA damage correlates with enhanced transcription of downstream p53 target genes. Low levels of phosphoserine-15 p53 are detectable within 6 hr after IR in AT cells, whereas lymphoblasts from normal individuals exhibit this modification within 1 hr. In contrast, phosphorylation of p53 on serine-15 is similar in normal and AT cells after UV irradiation. Our results indicate that p53 is phosphorylated in response to DNA damage, that this de novo phosphorylation may be involved in the subsequent induction and activation of p53, and that although ATM affects the kinetics of p53 phosphorylation after IR, it is not absolutely required for phosphorylation of p53 on serine-15.
Figures
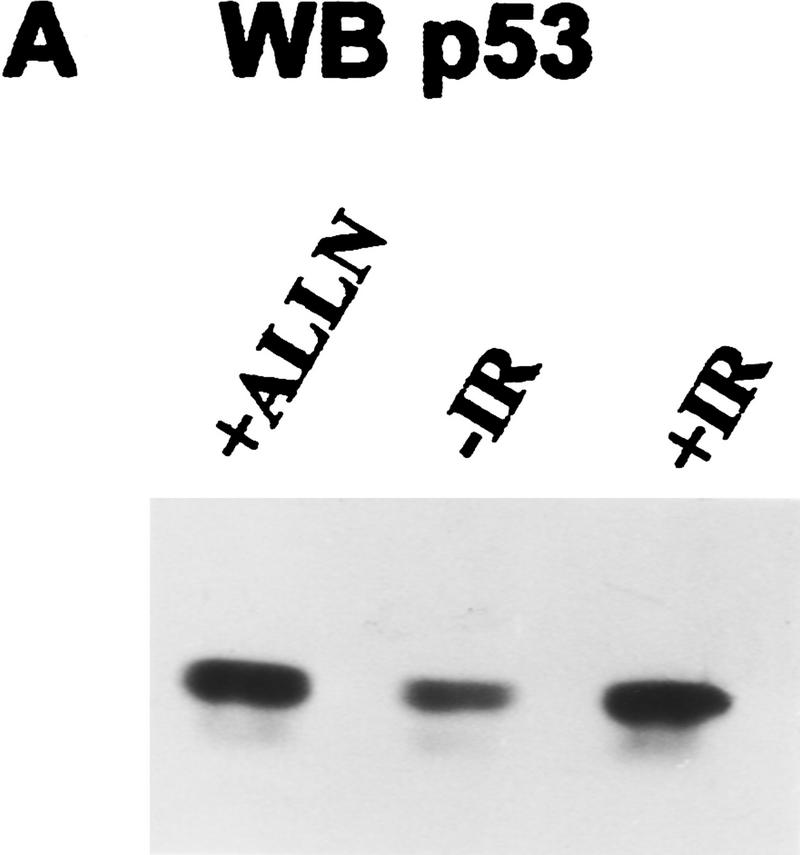
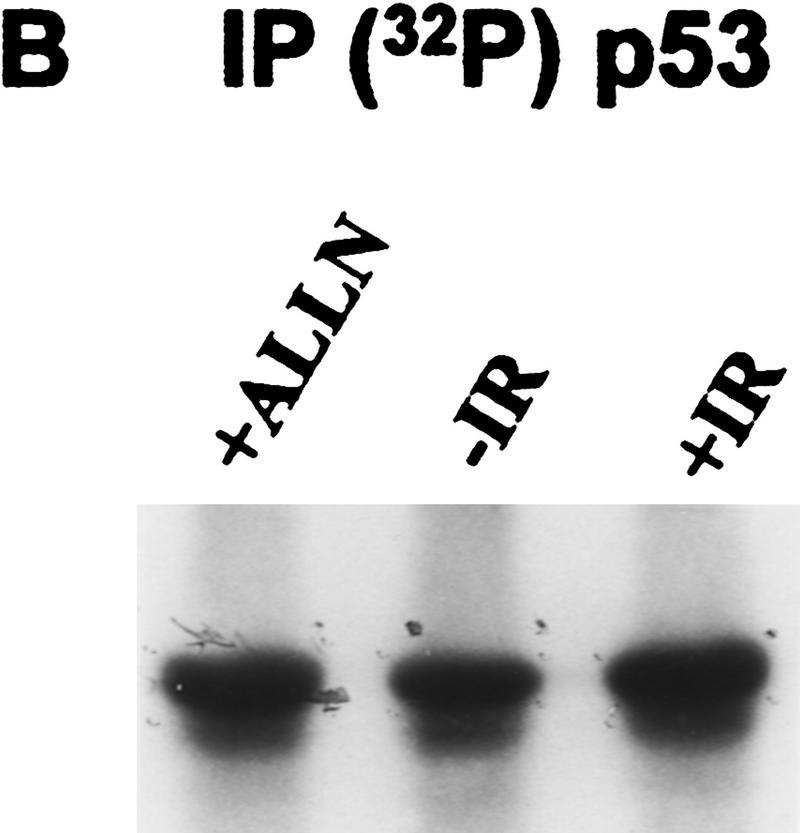
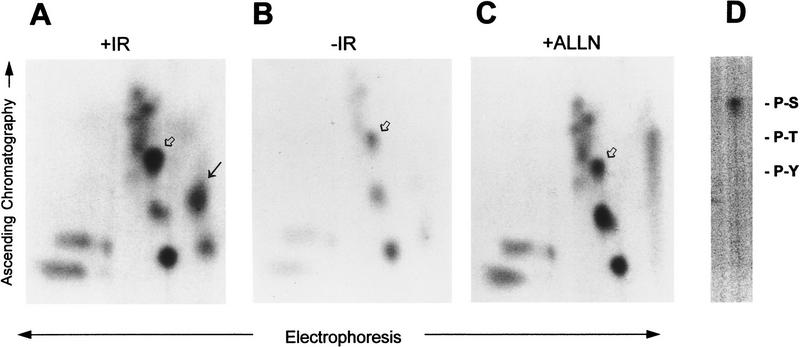
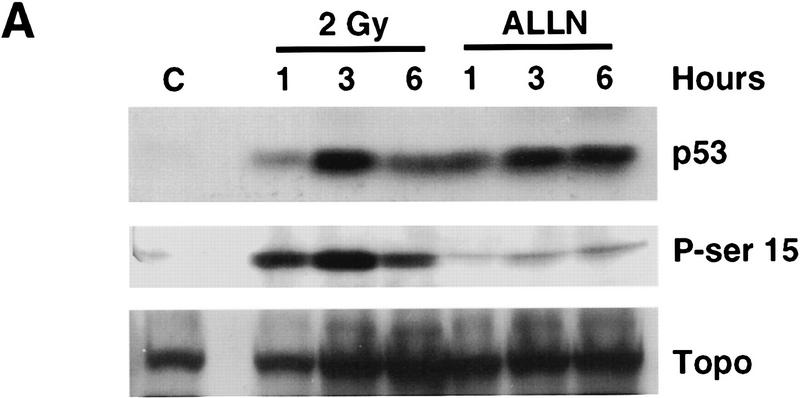
References
-
- Bogue MA, Zhu C, Aguilar-Cordova E, Donehower LA, Roth DB. p53 is required for both radiation-induced differentiation and rescue of V(D)J rearrangement in scid mouse thymocytes. Genes & Dev. 1996;10:553–565. - PubMed
-
- Boyle WJ, van der Geer P, Hunter T. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 1991;201:110–149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous