

The yeast adaptor protein complex, AP-3, is essential for the efficient delivery of alkaline phosphatase by the alternate pathway to the vacuole
- PMID: 9412470
- PMCID: PMC2132655
- DOI: 10.1083/jcb.139.7.1761
The yeast adaptor protein complex, AP-3, is essential for the efficient delivery of alkaline phosphatase by the alternate pathway to the vacuole
Abstract
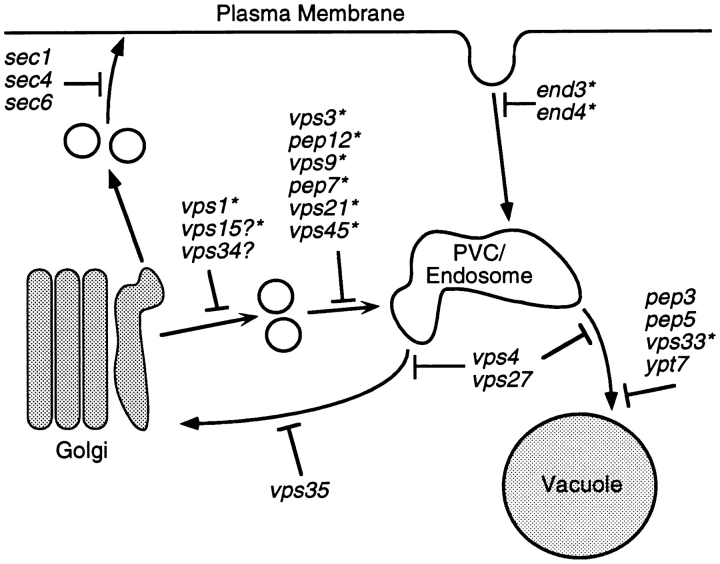
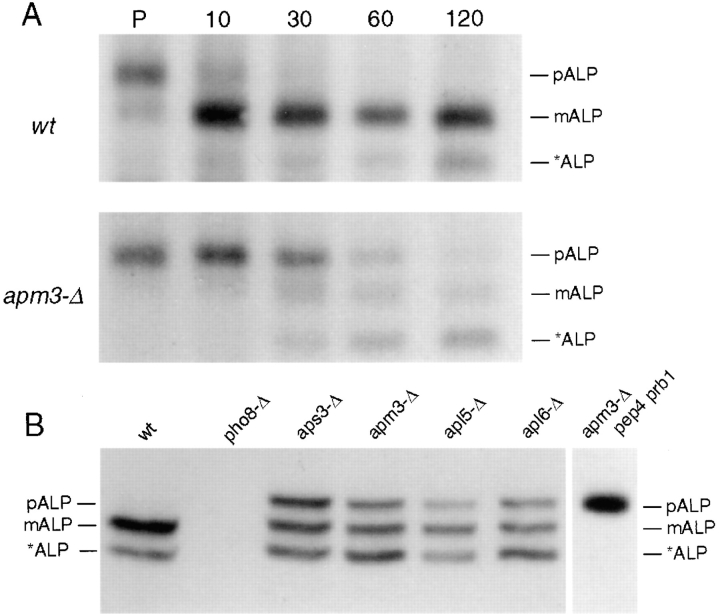
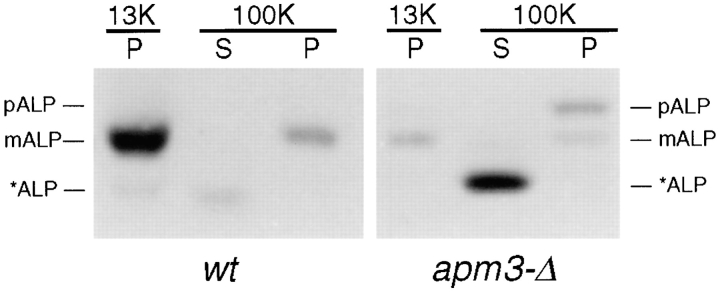
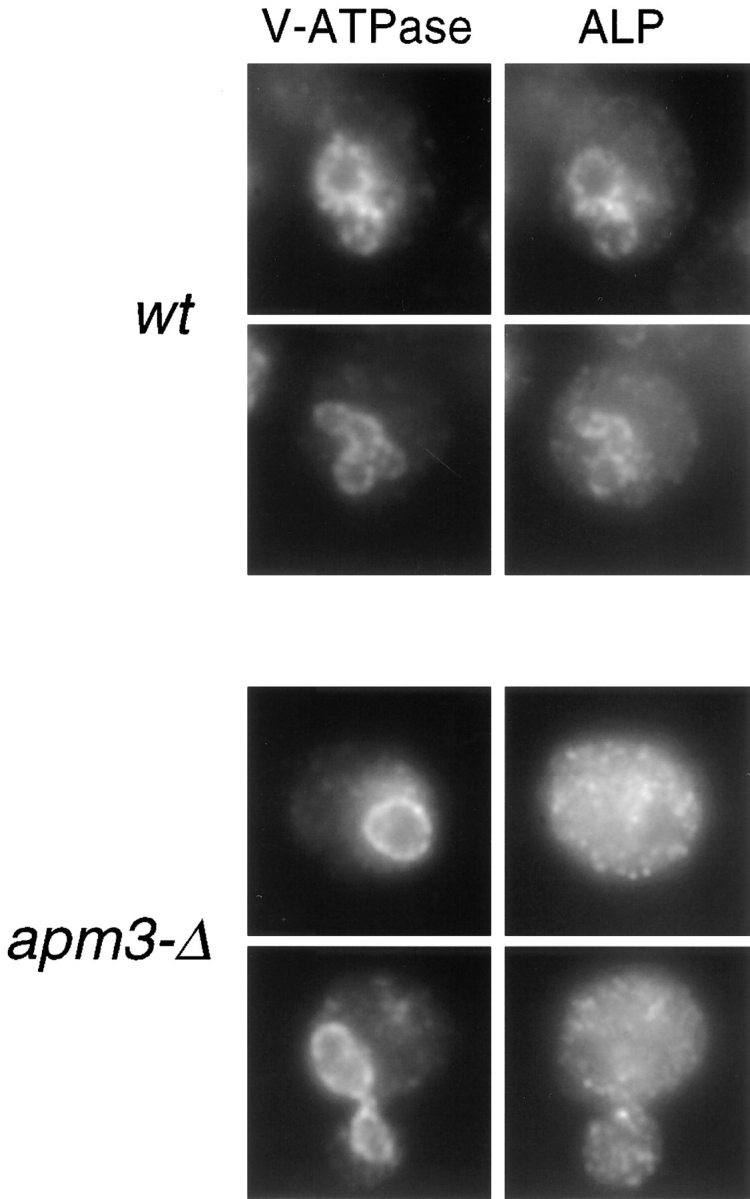
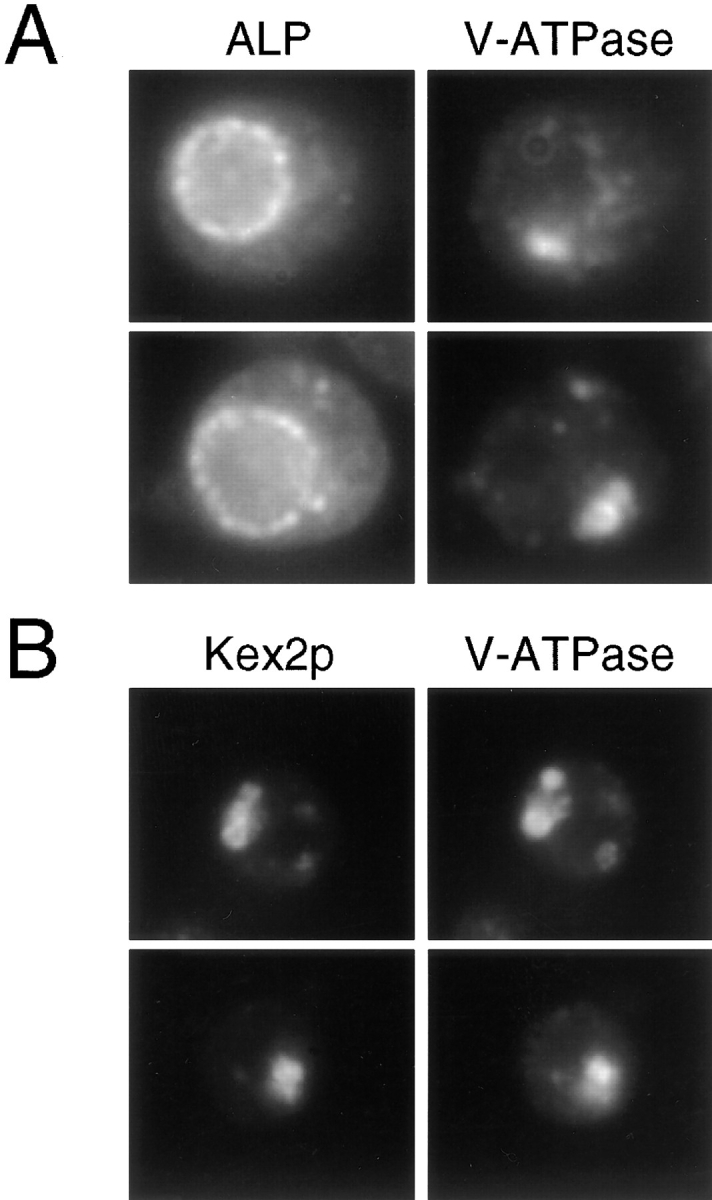
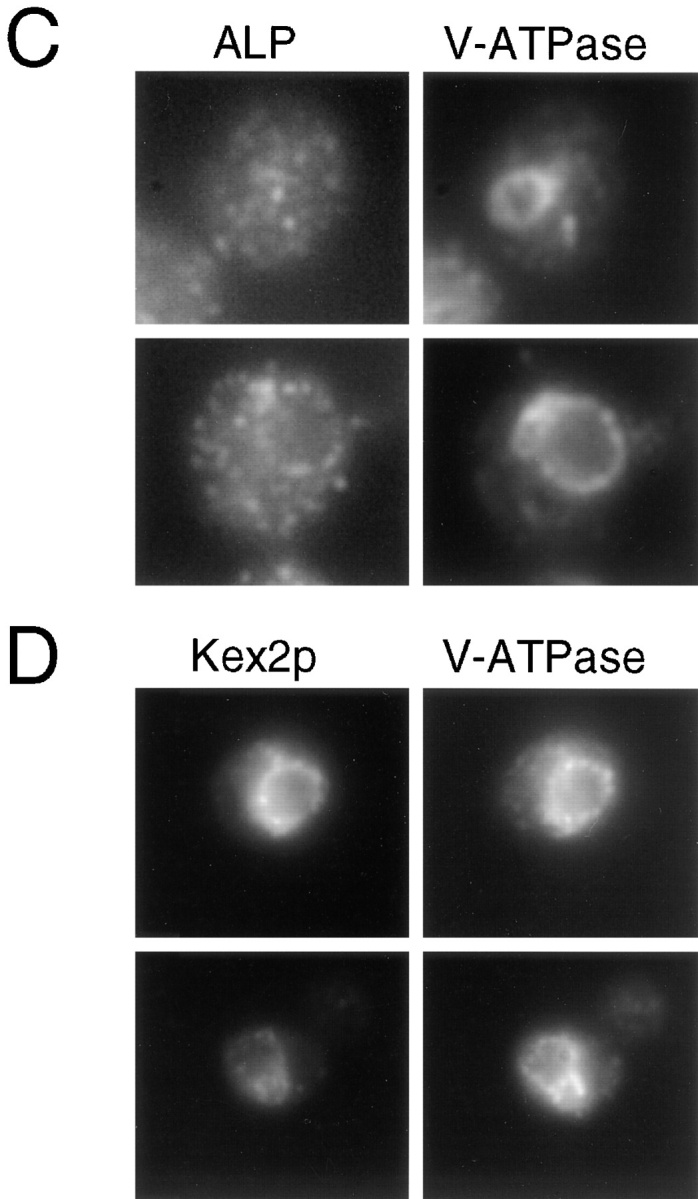
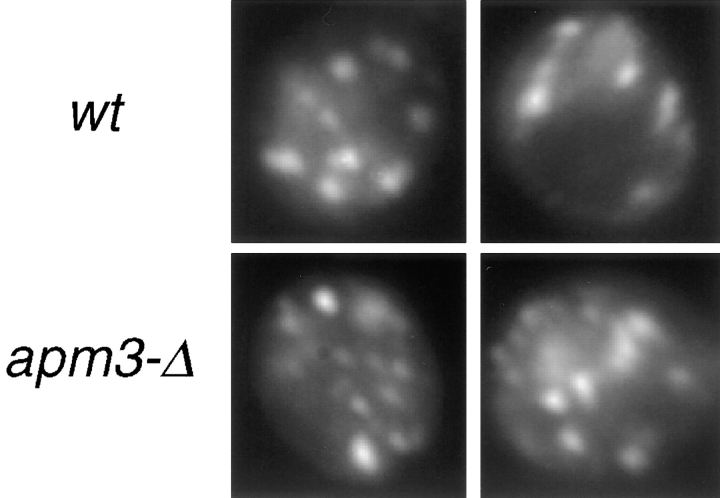
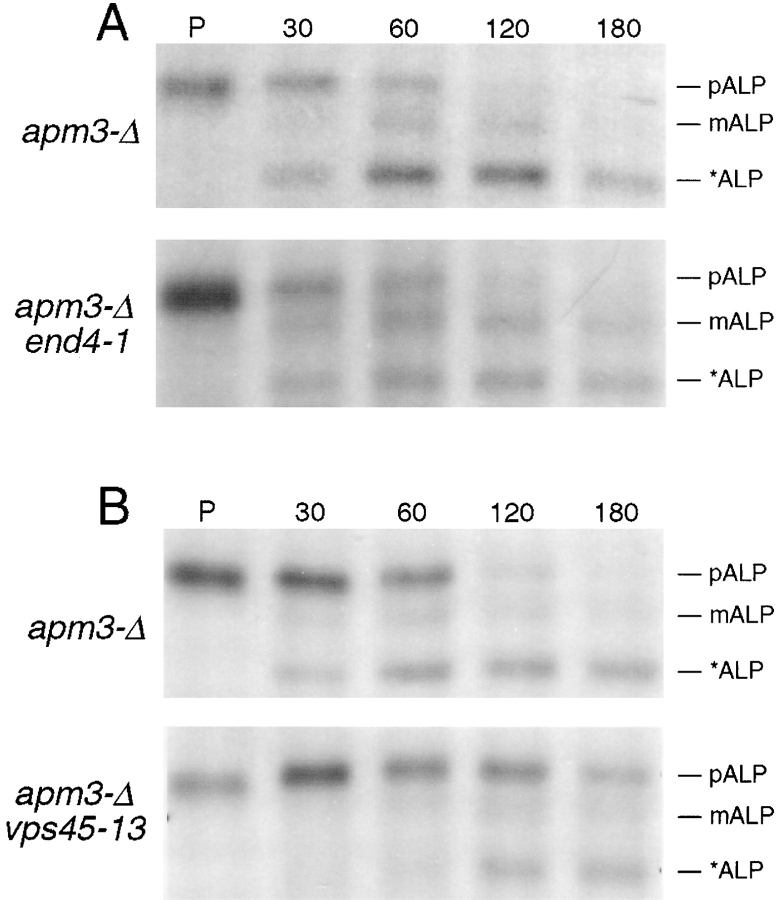
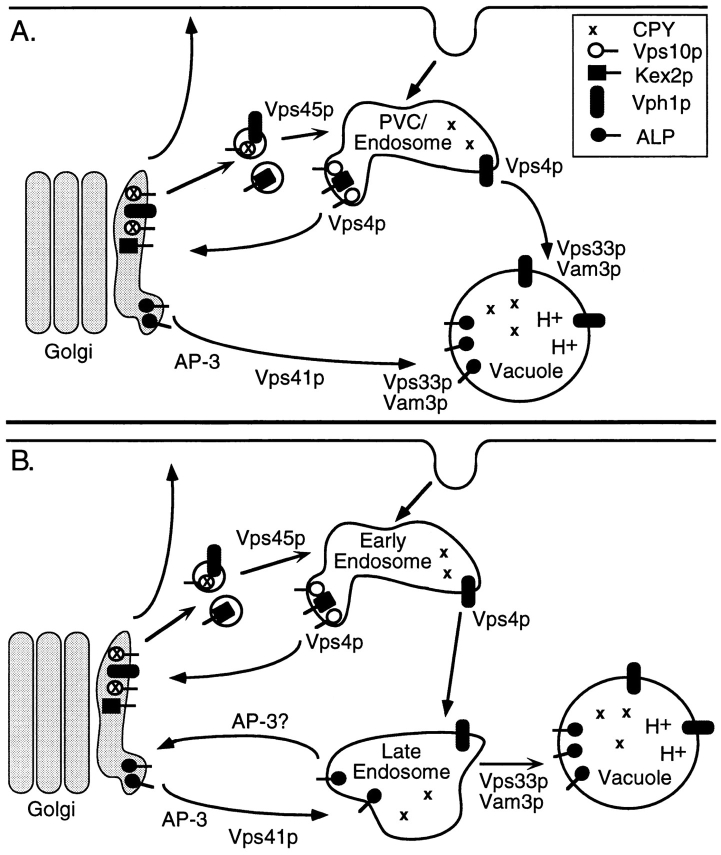
A novel clathrin adaptor-like complex, adaptor protein (AP)-3, has recently been described in yeast and in animals. To gain insight into the role of yeast AP-3, a genetic strategy was devised to isolate gene products that are required in the absence of the AP-3 mu chain encoded by APM3. One gene identified by this synthetic lethal screen was VPS45. The Vps pathway defines the route that several proteins, including carboxypeptidase Y, take from the late Golgi to the vacuole. However, vacuolar alkaline phosphatase (ALP) is transported via an alternate, intracellular route. This suggested that the apm3-Delta vps45 synthetic phenotype could be caused by a block in both the alternate and the Vps pathways. Here we demonstrate that loss of function of the AP-3 complex results in slowed processing and missorting of ALP. ALP is no longer localized to the vacuole membrane by immunofluorescence, but is found in small punctate structures throughout the cell. This pattern is distinct from the Golgi marker Kex2p, which is unaffected in AP-3 mutants. We also show that in the apm3-Delta mutant some ALP is delivered to the vacuole by diversion into the Vps pathway. Class E vps mutants accumulate an exaggerated prevacuolar compartment containing membrane proteins on their way to the vacuole or destined for recycling to the Golgi. Surprisingly, in AP-3 class E vps double mutants these proteins reappear on the vacuole. We suggest that some AP-3-dependent cargo proteins that regulate late steps in Golgi to vacuole transport are diverted into the Vps pathway allowing completion of transfer to the vacuole in the class E vps mutant.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
