

Potassium channel distribution, clustering, and function in remyelinating rat axons
- PMID: 9412484
- PMCID: PMC6793423
- DOI: 10.1523/JNEUROSCI.18-01-00036.1998
Potassium channel distribution, clustering, and function in remyelinating rat axons
Abstract
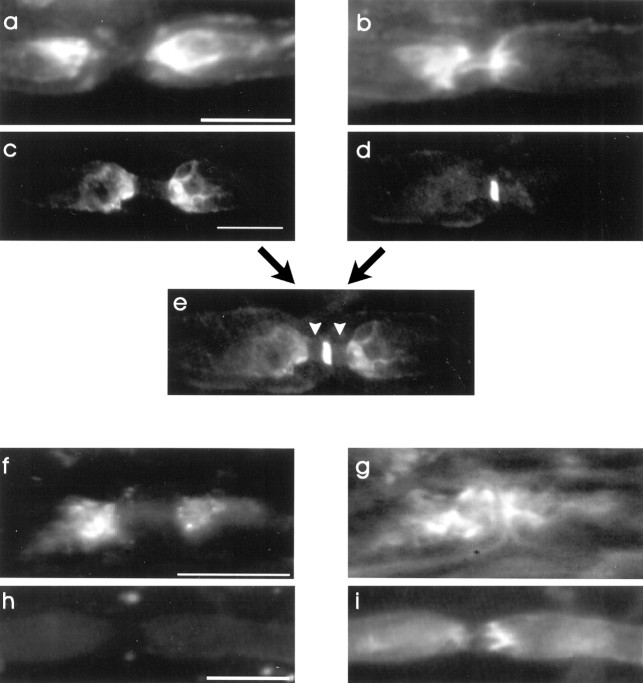
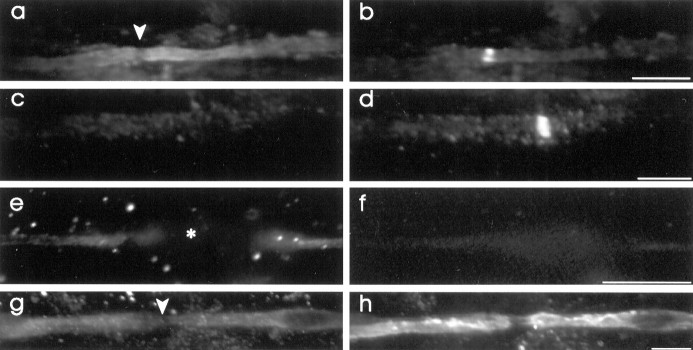
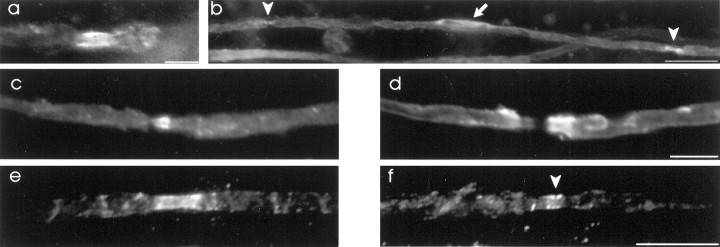
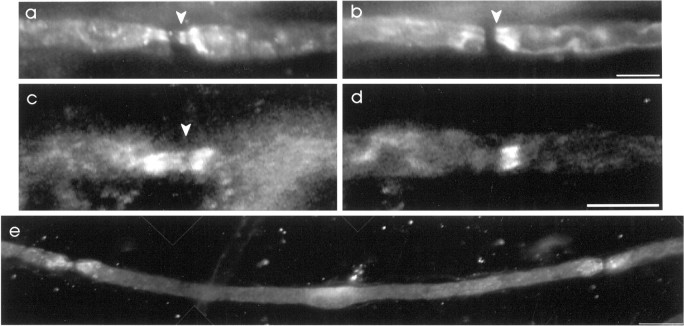
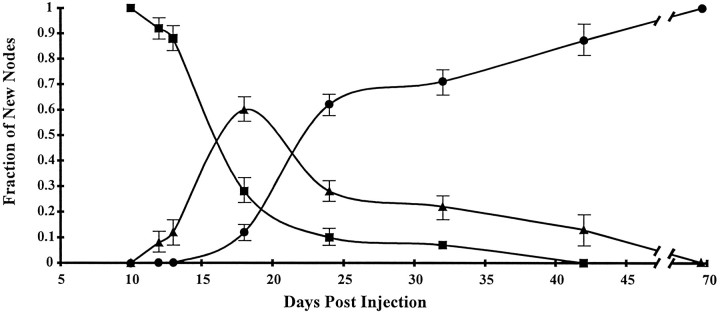
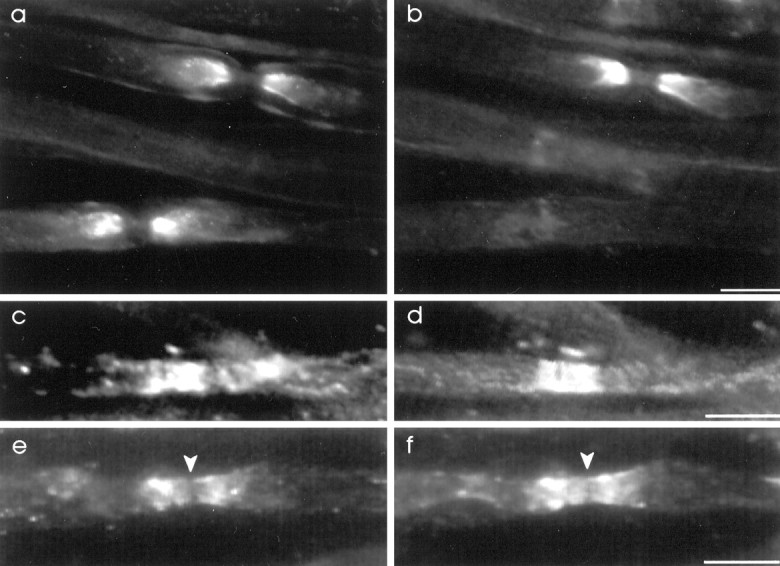
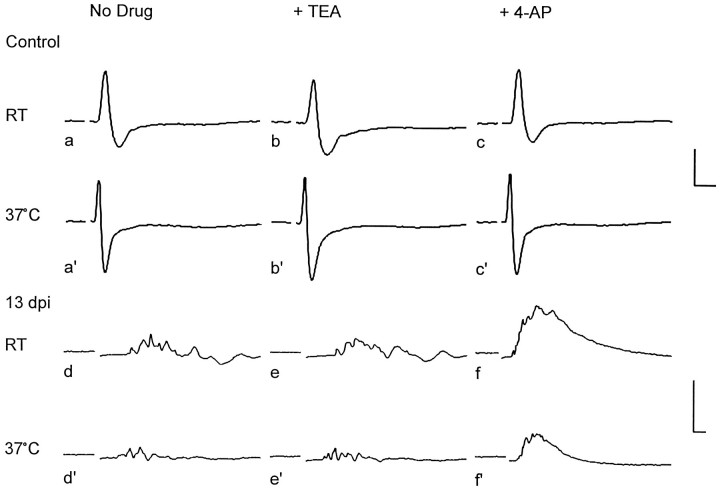
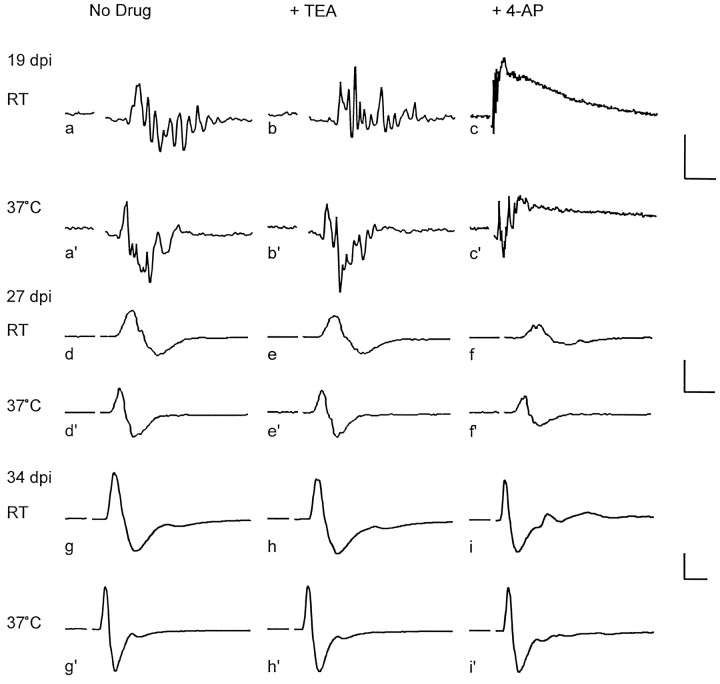
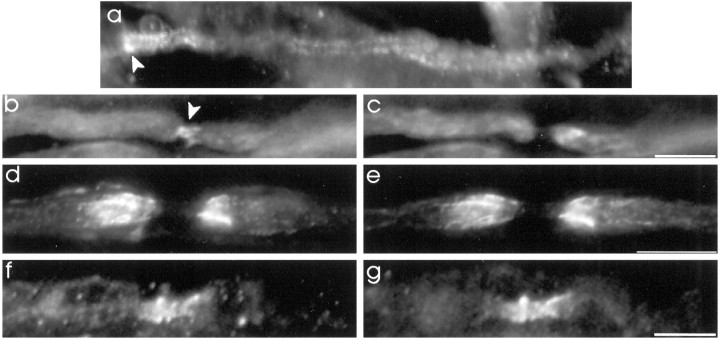
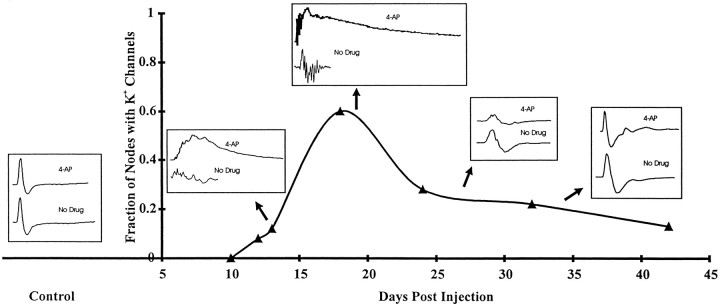
The K+ channel alpha-subunits Kv1.1 and Kv1.2 and the cytoplasmic beta-subunit Kvbeta2 were detected by immunofluorescence microscopy and found to be colocalized at juxtaparanodes in normal adult rat sciatic nerve. After demyelination by intraneural injection of lysolecithin, and during remyelination, the subcellular distributions of Kv1.1, Kv1.2, and Kvbeta2 were reorganized. At 6 d postinjection (dpi), axons were stripped of myelin, and K+ channels were found to be dispersed across zones that extended into both nodal and internodal regions; a few days later they were undetectable. By 10 dpi, remyelination was underway, but Kv1.1 immunoreactivity was absent at newly forming nodes of Ranvier. By 14 dpi, K+ channels were detected but were in the nodal gap between Schwann cells. By 19 dpi, most new nodes had Kv1.1, Kv1.2, and Kvbeta2, which precisely colocalized. However, this nodal distribution was transient. By 24 dpi, the majority of K+ channels was clustered within paranodal regions of remyelinated axons, leaving a gap that overlapped with Na+ channel immunoreactivity. Inhibition of Schwann cell proliferation delayed both remyelination and the development of the K+ channel distributions described. Conduction studies indicate that neither 4-aminopyridine (4-AP) nor tetraethylammonium alters normal nerve conduction. However, during remyelination, 4-AP profoundly increased both compound action potential amplitude and duration. The level of this effect matched closely the nodal presence of these voltage-dependent K+ channels. Our results suggest that K+ channels may have a significant effect on conduction during remyelination and that Schwann cells are important in K+ channel redistribution and clustering.
Figures
References
-
- Bekele-Arcuri Z, Matos MF, Manganas MM, Strassle BW, Monaghan MM, Rhodes KJ, Trimmer JS. Generation and characterization of subtype-specific monoclonal antibodies to K+ channel α- and β-subunit polypeptides. Neuropharmacology. 1996;35:851–865. - PubMed
-
- Castle NA, Fadous SR, Logothetis DE, Wang GK. 4-Aminopyridine binding and slow inactivation are mutually exclusive in rat Kv1.1 and Shaker potassium channels. Mol Pharmacol. 1994;46:1175–1181. - PubMed
-
- Chiu SY, Ritchie JM. Potassium channels in nodal and internodal axonal membrane of mammalian myelinated fibres. Nature. 1980;284:170–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous