

Peripheral target regulation of the development and survival of spinal sensory and motor neurons in the chick embryo
- PMID: 9412513
- PMCID: PMC6793405
- DOI: 10.1523/JNEUROSCI.18-01-00356.1998
Peripheral target regulation of the development and survival of spinal sensory and motor neurons in the chick embryo
Abstract
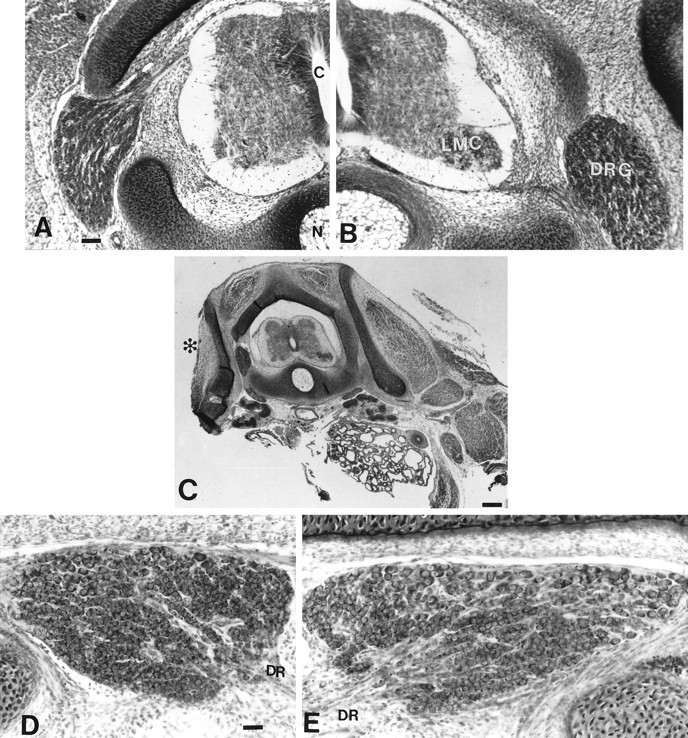
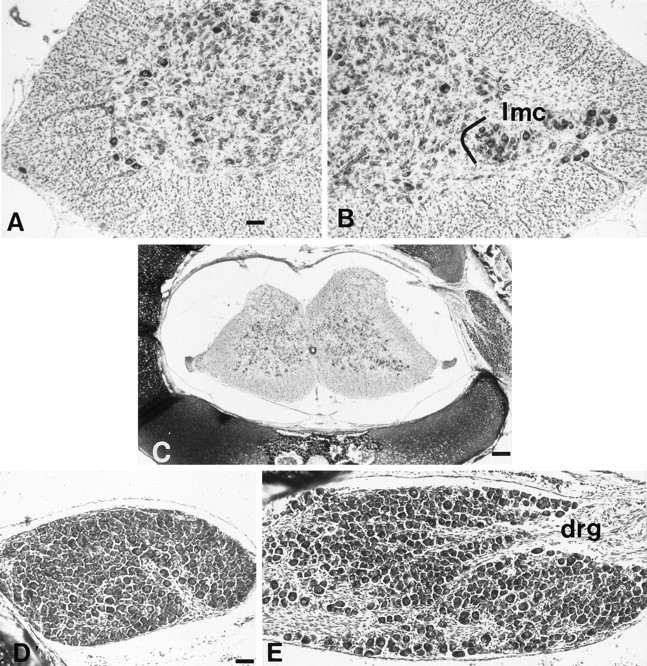
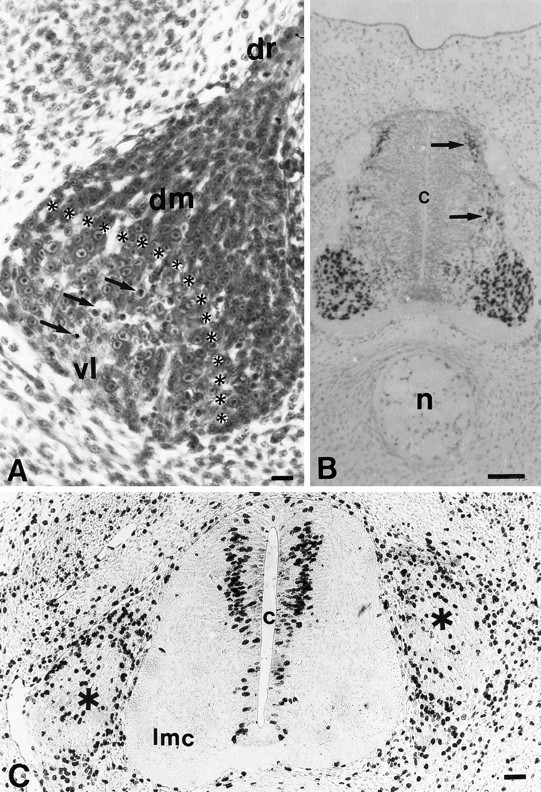
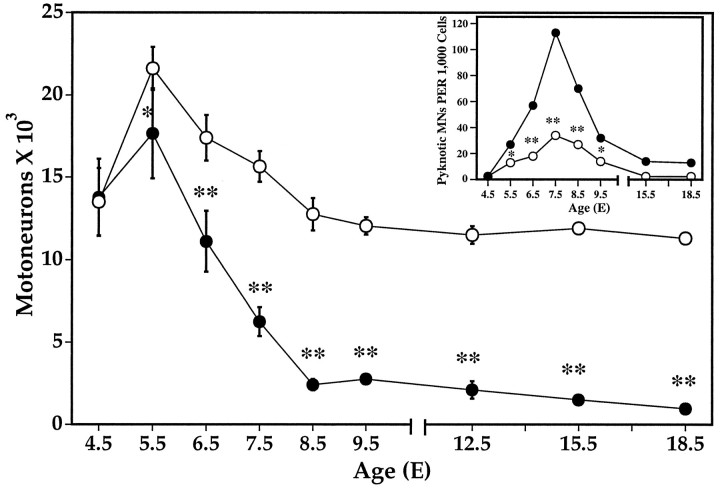
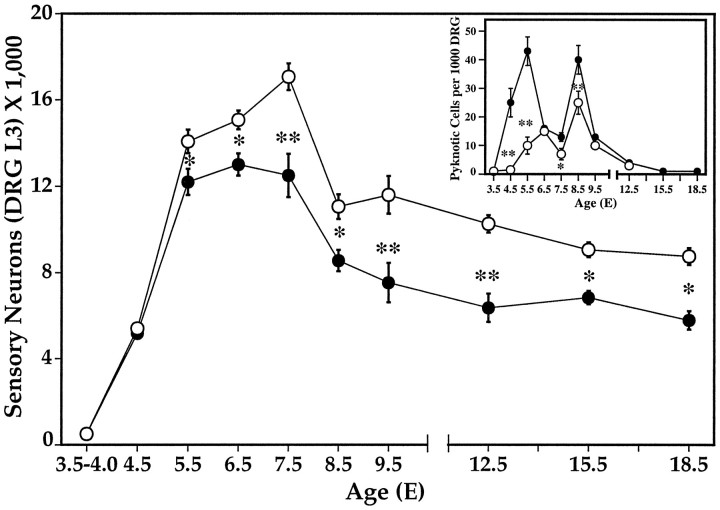
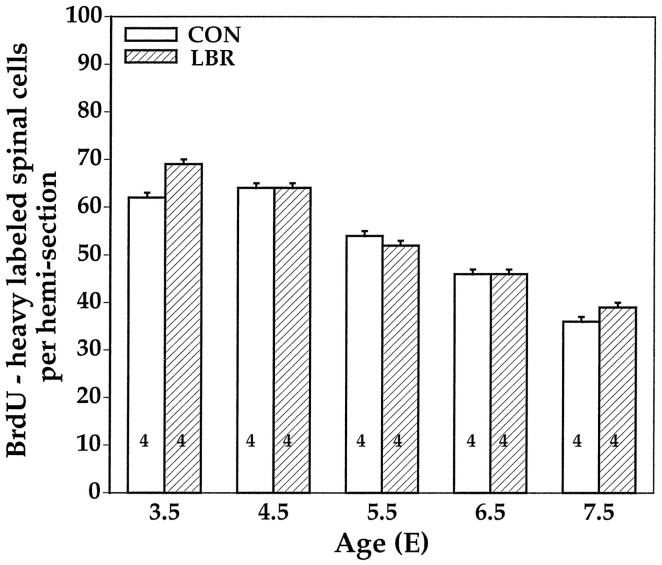
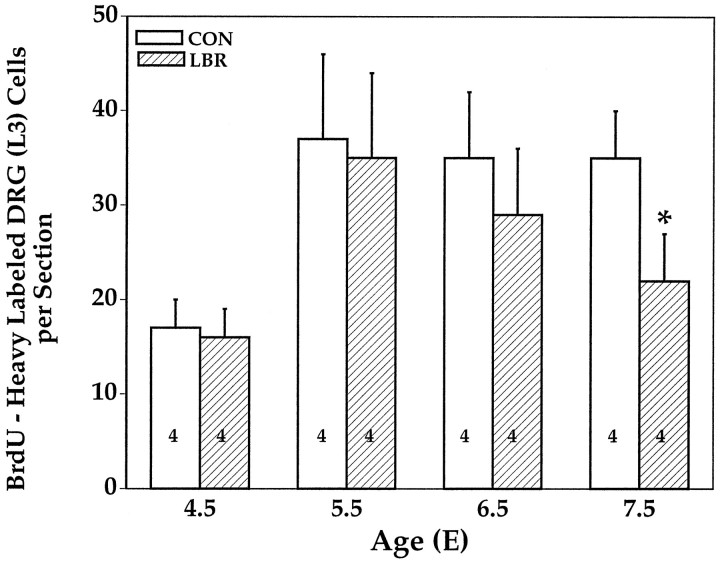
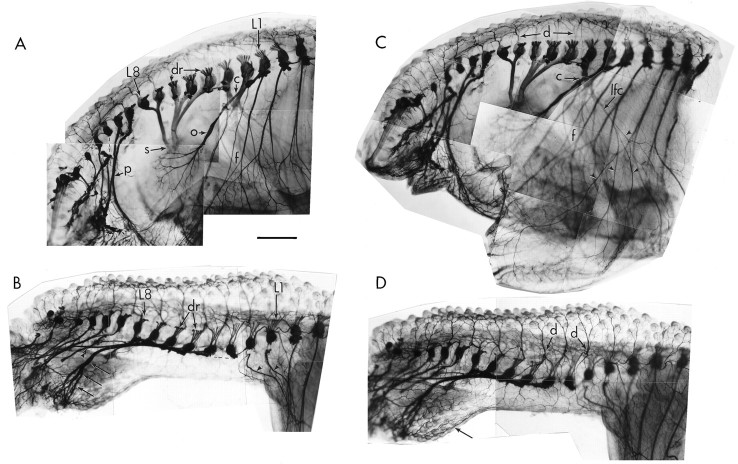
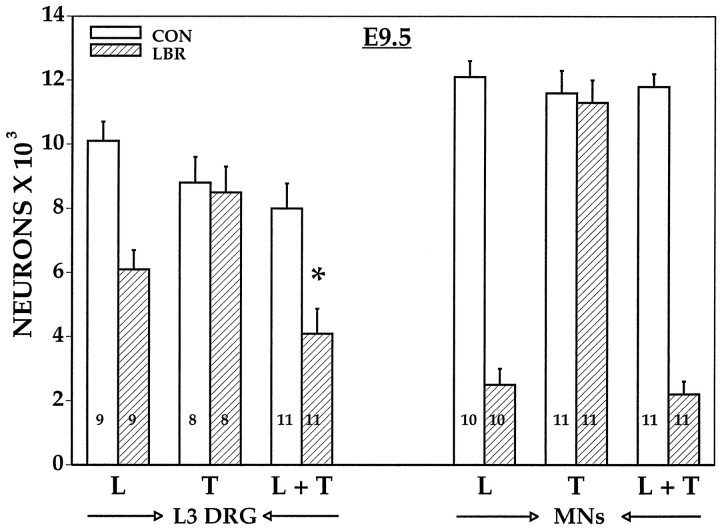
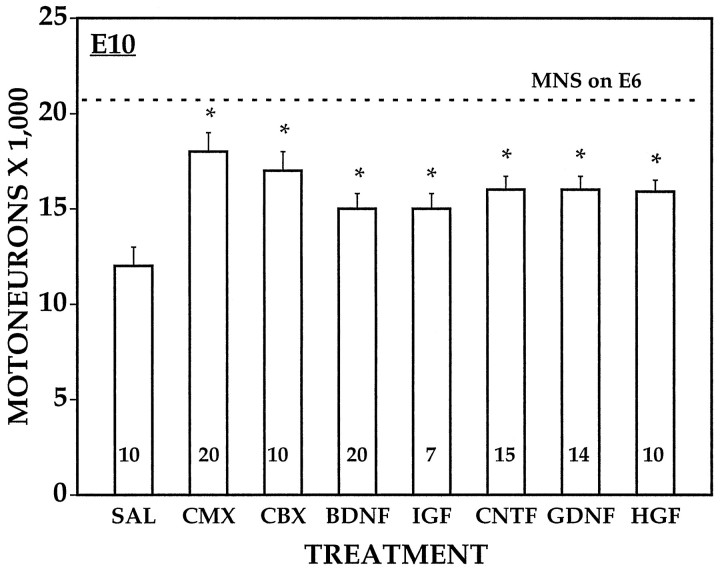
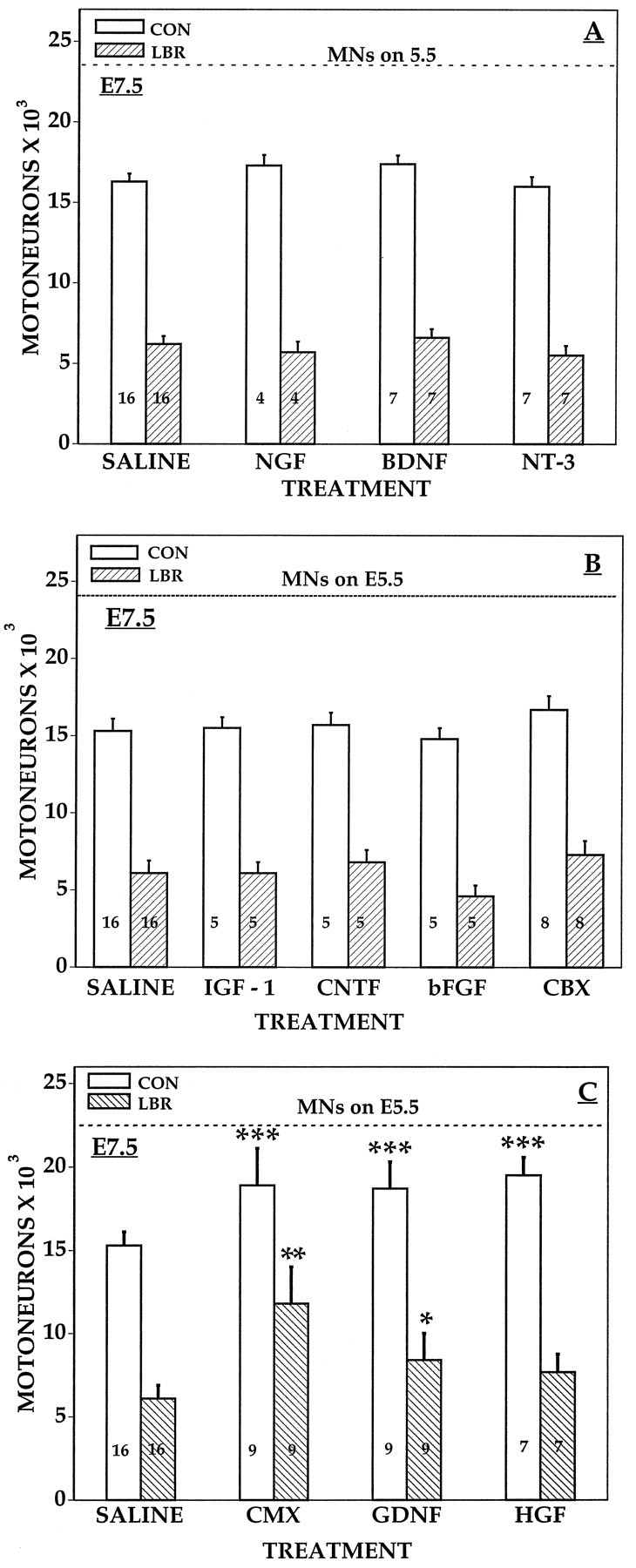
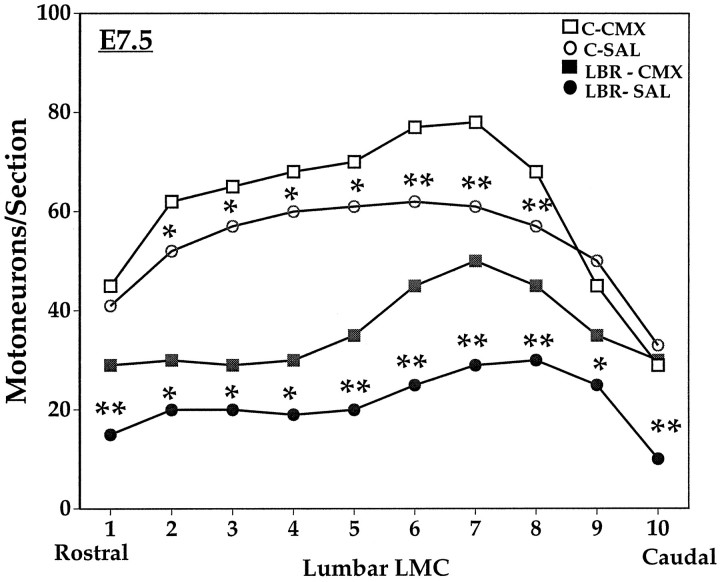
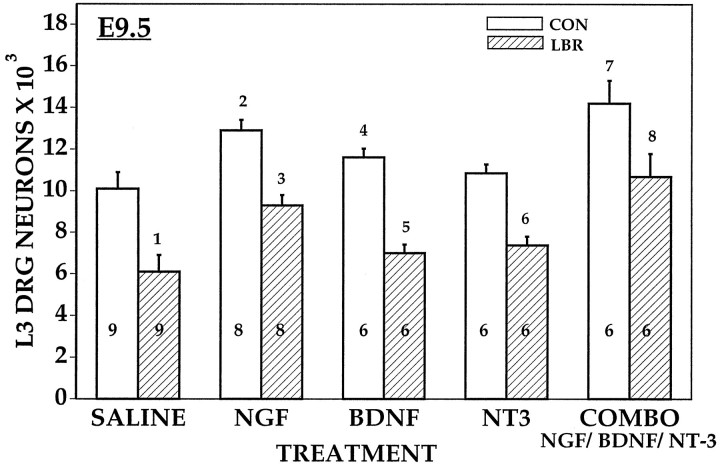
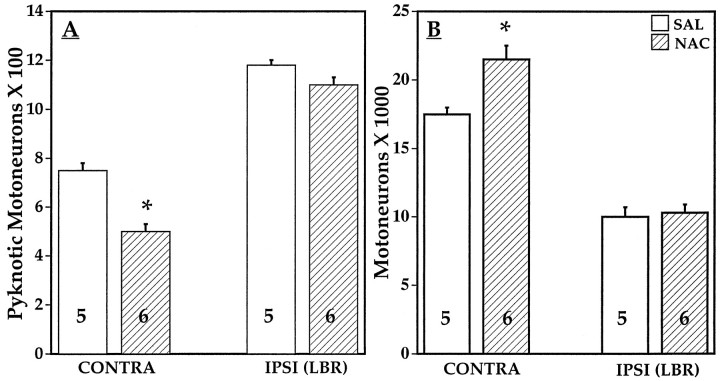
Unilateral limb-bud removal (LBR) before the outgrowth of sensory or motor neurons to the leg of chick embryos was used to examine the role of limb (target)-derived signals in the development and survival of lumbar motoneurons and sensory neurons in the dorsal root ganglia (DRG). After LBR, motor and sensory neurons underwent normal initial histological differentiation, and cell growth in both populations was unaffected. Before their death, target-deprived motoneurons also expressed a cell-specific marker, the homeodomain protein islet-1. Proliferation of sensory and motor precursor cells was also unaffected by LBR, and the migration of neural crest cells to the DRG and of motoneurons into the ventral horn occurred normally. During the normal period of programmed cell death (PCD), increased numbers of both sensory and motor neurons degenerated after LBR. However, whereas motoneuron loss increased by 40-50% (90% total), only approximately 25% more sensory neurons degenerated after LBR. A significant number of the surviving sensory neurons projected to aberrant targets in the tail after LBR, and many of these were lost after ablation of both the limb and tail. Treatment with neurotrophic factors (or muscle extract) rescued sensory and motor neurons from cell death after LBR without affecting precursor proliferation of either population. Activity blockade with curare failed to rescue motoneurons after LBR, and combined treatment with curare plus muscle extract was no more effective than muscle extract alone. Treatment with the antioxidant N-acetylcysteine rescued motoneurons from normal cell death but not after LBR. Two specific inhibitors of the interleukin beta1 converting enzyme (ICE) family of cysteine proteases also failed to prevent motoneuron death after LBR. Taken together these data provide definitive evidence that the loss of spinal neurons after LBR cannot be attributed to altered proliferation, migration, or differentiation. Rather, in the absence of limb-derived trophic signals, the affected neurons fail to survive and undergo PCD. Although normal cell death and cell death after target deprivation share many features in common, the intracellular pathways of cell death in the two may be distinct.
Figures
Similar articles
-
Neurotrophin-3 promotes the differentiation of muscle spindle afferents in the absence of peripheral targets.J Neurosci. 1997 Jun 1;17(11):4262-74. doi: 10.1523/JNEUROSCI.17-11-04262.1997. J Neurosci. 1997. PMID: 9151743 Free PMC article.
-
A novel type of programmed neuronal death in the cervical spinal cord of the chick embryo.J Neurosci. 1996 Jun 1;16(11):3685-703. doi: 10.1523/JNEUROSCI.16-11-03685.1996. J Neurosci. 1996. PMID: 8642412 Free PMC article.
-
Expression and regulation of brain-derived neurotrophic factor and neurotrophin-3 mRNAs in distinct avian motoneuron subsets.J Neurobiol. 1996 Mar;29(3):277-92. doi: 10.1002/(SICI)1097-4695(199603)29:3<277::AID-NEU1>3.0.CO;2-6. J Neurobiol. 1996. PMID: 8907158
-
Neurotrophic interactions in the development of spinal cord motoneurons.Ciba Found Symp. 1988;138:152-71. doi: 10.1002/9780470513675.ch10. Ciba Found Symp. 1988. PMID: 3058426 Review.
-
Spontaneous rhythmic activity in early chick spinal cord influences distinct motor axon pathfinding decisions.Brain Res Rev. 2008 Jan;57(1):77-85. doi: 10.1016/j.brainresrev.2007.06.021. Epub 2007 Aug 1. Brain Res Rev. 2008. PMID: 17920131 Free PMC article. Review.
Cited by
-
Effect of Strychinine, a Glycine Inhibitor, on the Programmed Cell Death of Motoneurons during the Chick Development.Exp Neurobiol. 2011 Dec;20(4):176-80. doi: 10.5607/en.2011.20.4.176. Epub 2011 Dec 29. Exp Neurobiol. 2011. PMID: 22355262 Free PMC article.
-
Intrinsic control of neuronal diversity and synaptic specificity in a proprioceptive circuit.Elife. 2020 Aug 18;9:e56374. doi: 10.7554/eLife.56374. Elife. 2020. PMID: 32808924 Free PMC article.
-
An early broad competence of motoneurons to express ER81 is later sculpted by the periphery.J Neurosci. 2004 Nov 3;24(44):9789-98. doi: 10.1523/JNEUROSCI.3409-04.2004. J Neurosci. 2004. PMID: 15525764 Free PMC article.
-
Thioredoxin-2 Modulates Neuronal Programmed Cell Death in the Embryonic Chick Spinal Cord in Basal and Target-Deprived Conditions.PLoS One. 2015 Nov 5;10(11):e0142280. doi: 10.1371/journal.pone.0142280. eCollection 2015. PLoS One. 2015. PMID: 26540198 Free PMC article.
-
Inhibition of electrical activity by retroviral infection with Kir2.1 transgenes disrupts electrical differentiation of motoneurons.PLoS One. 2008 Aug 13;3(8):e2971. doi: 10.1371/journal.pone.0002971. PLoS One. 2008. PMID: 18698433 Free PMC article.
References
-
- Acheson A, Connover JC, Fandi JP, Dechiara TM, Russell M, Thadani A, Squinto SP, Yancopoulas GD, Lindsay RM. A BDNF autocrine loop in adult sensory neurons prevents cell death. Nature. 1995;374:450–453. - PubMed
-
- Bartlett SE, Reynolds AJ, Weible M, Haydon K, Hendry IA. In sympathetic but not sensory neurons PI-3 kinase is important for NGF-dependent survival and the retrograde transport of 125I-βNGF. Brain Res. 1997;761:257–262. - PubMed
-
- Bueker ED. Intracentral and peripheral factors in the differentiation of motor neurons in transplanted lumbo-sacral spinal cords of chick embryos. J Exp Zool. 1943;93:99–129.
-
- Bueker ED. Limb ablation experiments on the embryonic chick and its effect as observed on the mature nervous system. Anat Rec. 1947;97:157–174. - PubMed
-
- Burek MJ, Prevette D, Oppenheim RW. A temporal relationship between motoneuron birth date and onset of programmed cell death. Soc Neurosci Abstr. 1996;22:566.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources