

RNA recognition motif 2 of yeast Pab1p is required for its functional interaction with eukaryotic translation initiation factor 4G
- PMID: 9418852
- PMCID: PMC121449
- DOI: 10.1128/MCB.18.1.51
RNA recognition motif 2 of yeast Pab1p is required for its functional interaction with eukaryotic translation initiation factor 4G
Abstract
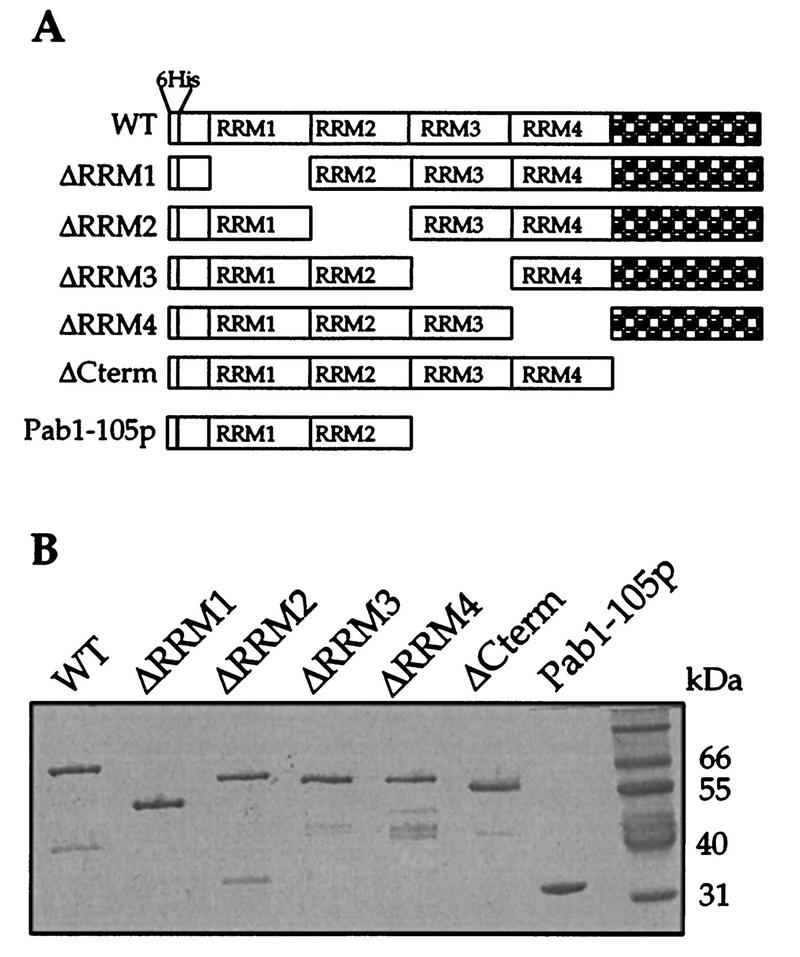
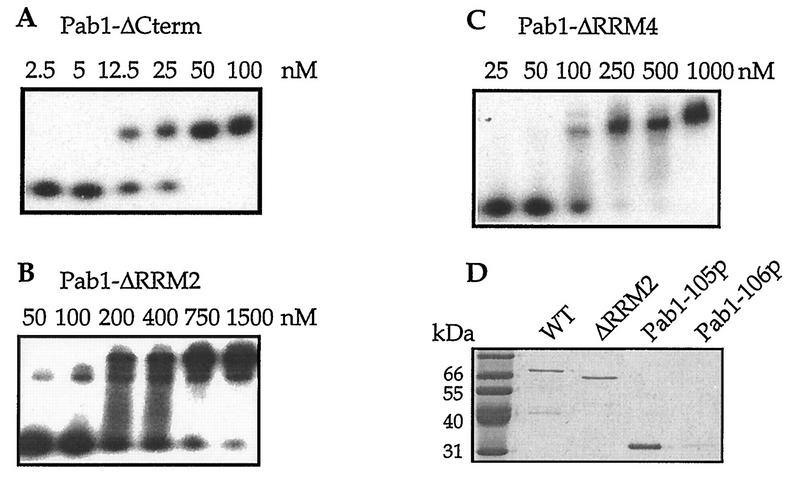
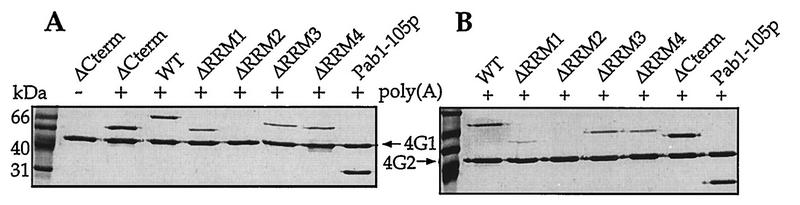
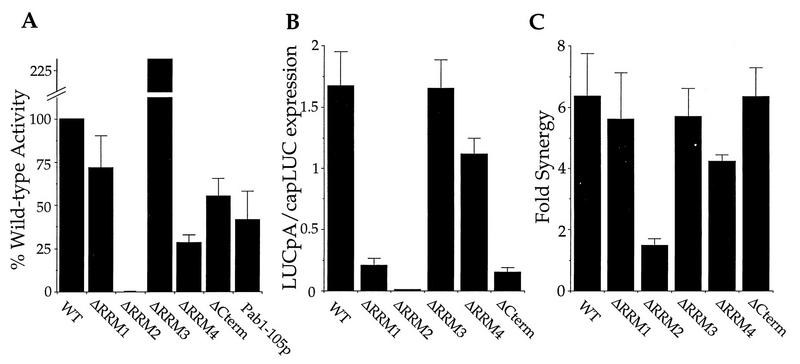
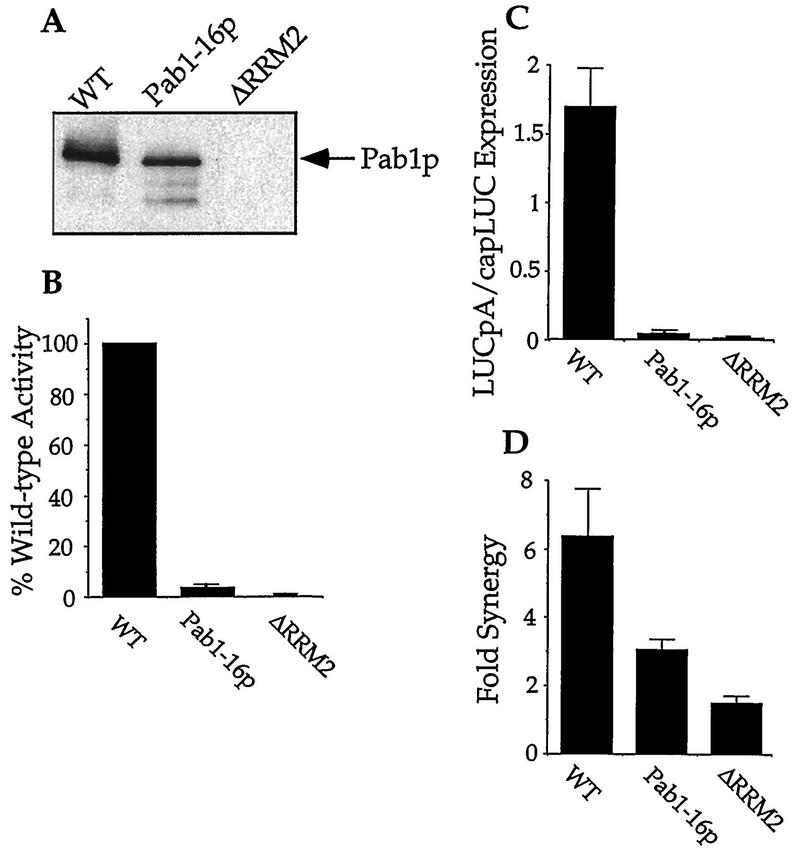
The eukaryotic mRNA 3' poly(A) tail and its associated poly(A)-binding protein (Pab1p) are important regulators of gene expression. One role for this complex in the yeast Saccharomyces cerevisiae is in translation initiation through an interaction with a 115-amino-acid region of the translation initiation factor eIF4G. The eIF4G-interacting domain of Pab1p was mapped to its second RNA recognition motif (RRM2) in an in vitro binding assay. Moreover, RRM2 of Pab1p was required for poly(A) tail-dependent translation in yeast extracts. An analysis of a site-directed Pab1p mutation which bound to eIF4G but did not stimulate translation of uncapped, polyadenylated mRNA suggested additional Pab1p-dependent events during translation initiation. These results support the model that the association of RRM2 of yeast Pab1p with eIF4G is a prerequisite for the poly(A) tail to stimulate the translation of mRNA in vitro.
Figures
Similar articles
-
The yeast poly(A)-binding protein Pab1p stimulates in vitro poly(A)-dependent and cap-dependent translation by distinct mechanisms.EMBO J. 1999 Jun 1;18(11):3153-63. doi: 10.1093/emboj/18.11.3153. EMBO J. 1999. PMID: 10357826 Free PMC article.
-
Binding of eukaryotic translation initiation factor 4E (eIF4E) to eIF4G represses translation of uncapped mRNA.Mol Cell Biol. 1997 Dec;17(12):6876-86. doi: 10.1128/MCB.17.12.6876. Mol Cell Biol. 1997. PMID: 9372919 Free PMC article.
-
Translation initiation factor eIF4G mediates in vitro poly(A) tail-dependent translation.Proc Natl Acad Sci U S A. 1997 Aug 19;94(17):9046-51. doi: 10.1073/pnas.94.17.9046. Proc Natl Acad Sci U S A. 1997. PMID: 9256432 Free PMC article.
-
Eukaryotic translation initiation: there are (at least) two sides to every story.Nat Struct Biol. 2000 May;7(5):356-61. doi: 10.1038/75120. Nat Struct Biol. 2000. PMID: 10802729 Review.
-
eIF4G-an integrator of mRNA metabolism?FEMS Yeast Res. 2016 Nov;16(7):fow087. doi: 10.1093/femsyr/fow087. Epub 2016 Sep 29. FEMS Yeast Res. 2016. PMID: 27694156 Review.
Cited by
-
Mass spectrometric identification of proteins that interact through specific domains of the poly(A) binding protein.Mol Genet Genomics. 2012 Sep;287(9):711-730. doi: 10.1007/s00438-012-0709-5. Epub 2012 Jul 27. Mol Genet Genomics. 2012. PMID: 22836166 Free PMC article.
-
Poly(A)-binding proteins regulate both mRNA deadenylation and decapping in yeast cytoplasmic extracts.RNA. 2001 Oct;7(10):1416-24. RNA. 2001. PMID: 11680846 Free PMC article.
-
CAF1 plays an important role in mRNA deadenylation separate from its contact to CCR4.Nucleic Acids Res. 2007;35(9):3002-15. doi: 10.1093/nar/gkm196. Epub 2007 Apr 16. Nucleic Acids Res. 2007. PMID: 17439972 Free PMC article.
-
Efficient cleavage of ribosome-associated poly(A)-binding protein by enterovirus 3C protease.J Virol. 2002 Mar;76(5):2062-74. doi: 10.1128/jvi.76.5.2062-2074.2002. J Virol. 2002. PMID: 11836384 Free PMC article.
-
A specific role for the C-terminal region of the Poly(A)-binding protein in mRNA decay.Nucleic Acids Res. 2007;35(18):6017-28. doi: 10.1093/nar/gkm452. Epub 2007 Aug 30. Nucleic Acids Res. 2007. PMID: 17766253 Free PMC article.
References
-
- Deardorff J A, Sachs A B. Differential effects of aromatic and charged residue substitutions in the RNA binding domains of the yeast poly(A)-binding protein. J Mol Biol. 1997;269:67–81. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous