

Multiple roles for the MyoD basic region in transmission of transcriptional activation signals and interaction with MEF2
- PMID: 9418854
- PMCID: PMC121453
- DOI: 10.1128/MCB.18.1.69
Multiple roles for the MyoD basic region in transmission of transcriptional activation signals and interaction with MEF2
Abstract
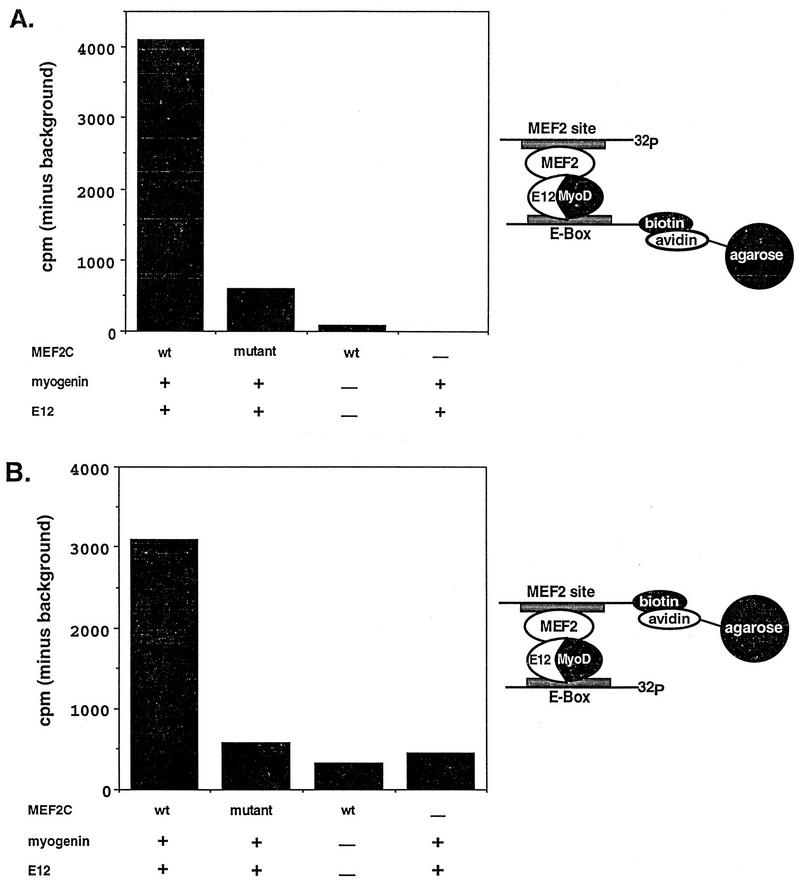
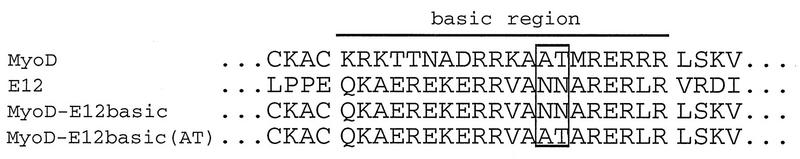
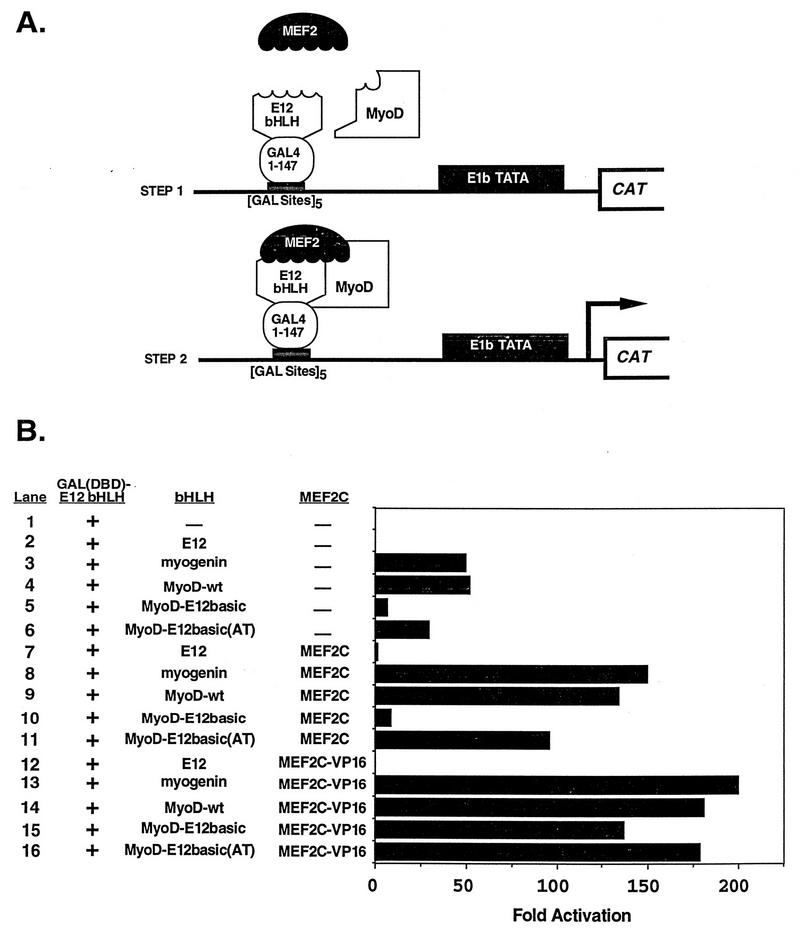
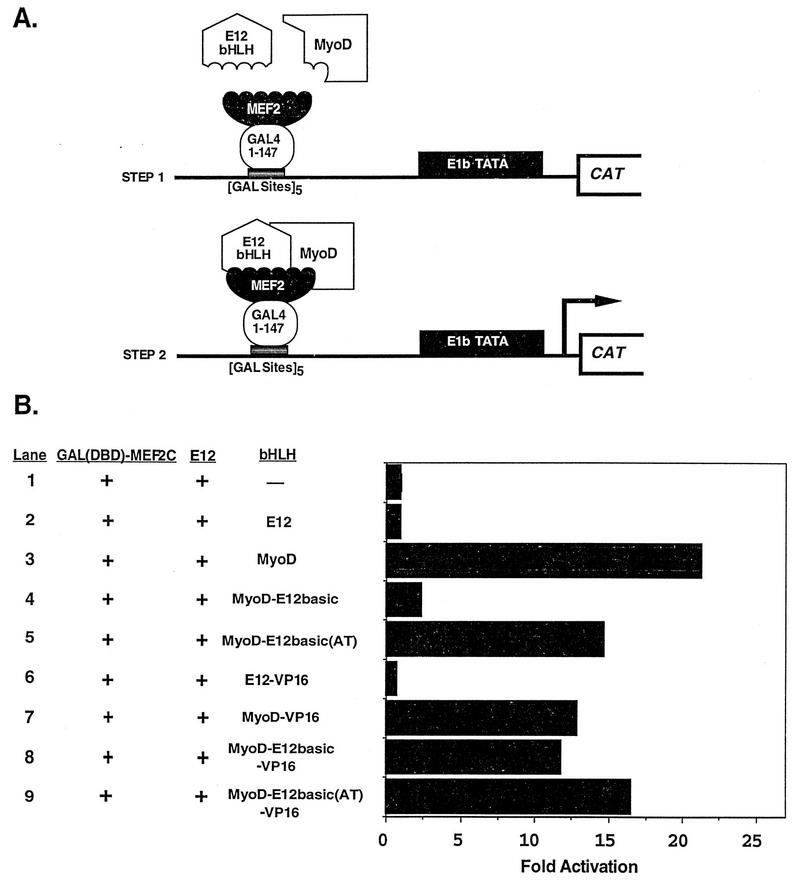
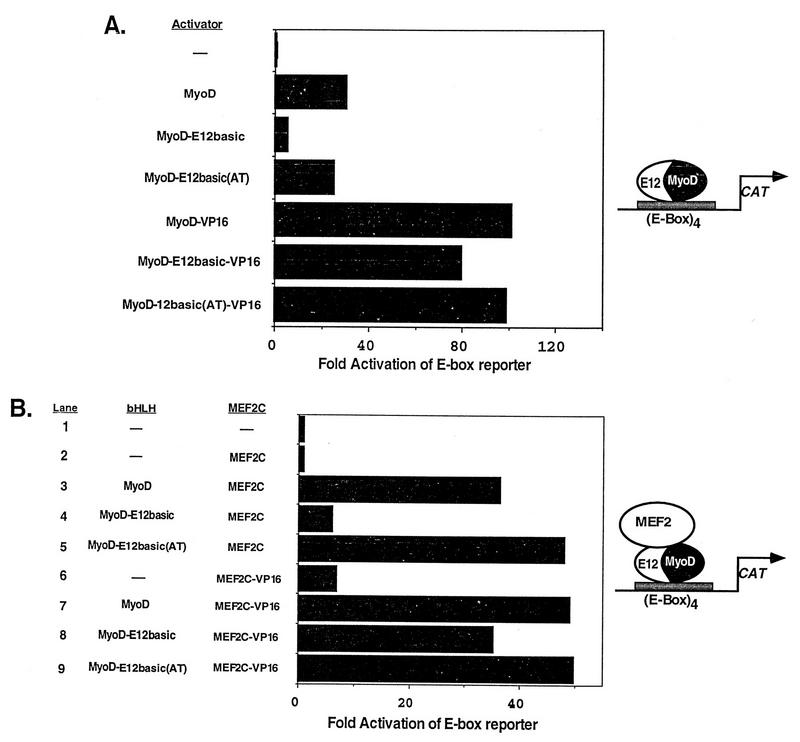
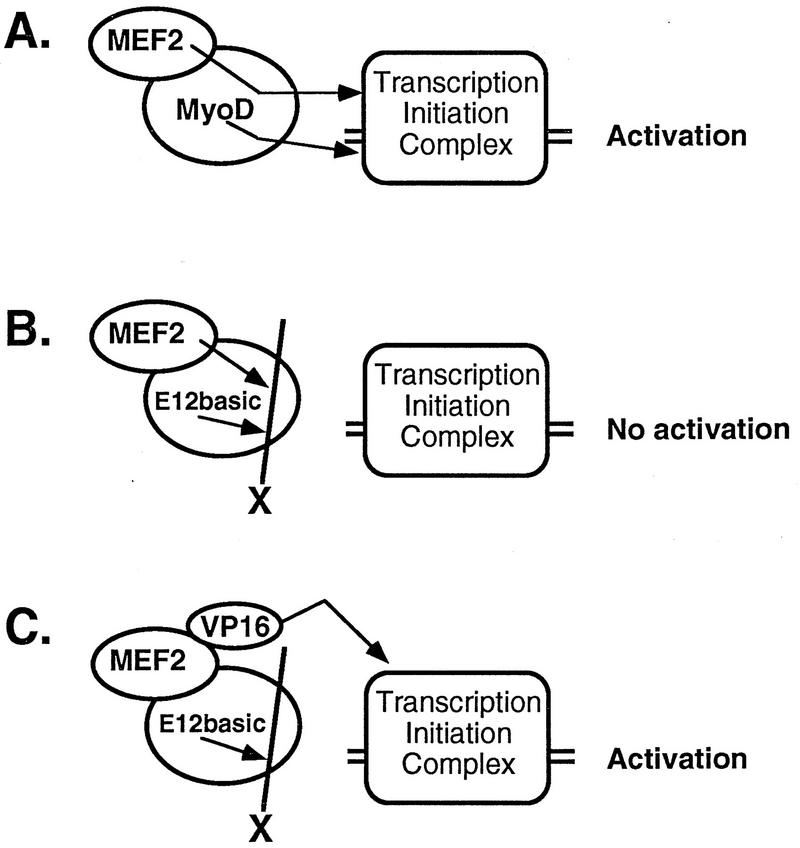
Establishment of skeletal muscle lineages is controlled by the MyoD family of basic helix-loop-helix (bHLH) transcription factors. The ability of these factors to initiate myogenesis is dependent on two conserved amino acid residues, alanine and threonine, in the basic domains of these factors. It has been postulated that these two residues may be responsible for the initiation of myogenesis via interaction with an essential myogenic cofactor. The myogenic bHLH proteins cooperatively activate transcription and myogenesis through protein-protein interactions with members of the myocyte enhancer factor 2 (MEF2) family of MADS domain transcription factors. MyoD-E12 heterodimers interact with MEF2 proteins to synergistically activate myogenesis, while homodimers of E12, which lack the conserved alanine and threonine residues in the basic domain, do not interact with MEF2. We have examined whether the myogenic alanine and threonine in the MyoD basic region are required for interaction with MEF2. Here, we show that substitution of the MyoD basic domain with that of E12 does not prevent interaction with MEF2. Instead, the inability of alanine-threonine mutants of MyoD to initiate myogenesis is due to a failure to transmit transcriptional activation signals provided either from the MyoD or the MEF2 activation domain. This defect in transcriptional transmission can be overcome by substitution of the MyoD or the MEF2 activation domain with the VP16 activation domain. These results demonstrate that myogenic bHLH-MEF2 interaction can be uncoupled from transcriptional activation and support the idea that the myogenic residues in myogenic bHLH proteins are essential for transmission of a transcriptional activation signal.
Figures
Similar articles
-
The basic domain of myogenic basic helix-loop-helix (bHLH) proteins is the novel target for direct inhibition by another bHLH protein, Twist.Mol Cell Biol. 1997 Nov;17(11):6563-73. doi: 10.1128/MCB.17.11.6563. Mol Cell Biol. 1997. PMID: 9343420 Free PMC article.
-
Molecular mechanisms of myogenic coactivation by p300: direct interaction with the activation domain of MyoD and with the MADS box of MEF2C.Mol Cell Biol. 1997 Feb;17(2):1010-26. doi: 10.1128/MCB.17.2.1010. Mol Cell Biol. 1997. PMID: 9001254 Free PMC article.
-
mef2c is activated directly by myogenic basic helix-loop-helix proteins during skeletal muscle development in vivo.Mech Dev. 2003 Sep;120(9):1021-32. doi: 10.1016/s0925-4773(03)00178-3. Mech Dev. 2003. PMID: 14550531
-
Combinatorial control of muscle development by basic helix-loop-helix and MADS-box transcription factors.Proc Natl Acad Sci U S A. 1996 Sep 3;93(18):9366-73. doi: 10.1073/pnas.93.18.9366. Proc Natl Acad Sci U S A. 1996. PMID: 8790335 Free PMC article. Review.
-
Muscle differentiation: more complexity to the network of myogenic regulators.Curr Opin Genet Dev. 1998 Oct;8(5):539-44. doi: 10.1016/s0959-437x(98)80008-7. Curr Opin Genet Dev. 1998. PMID: 9794824 Review.
Cited by
-
Histone Lysine Methylation and Long Non-Coding RNA: The New Target Players in Skeletal Muscle Cell Regeneration.Front Cell Dev Biol. 2021 Dec 3;9:759237. doi: 10.3389/fcell.2021.759237. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34926450 Free PMC article. Review.
-
ERK5 promotes Src-induced podosome formation by limiting Rho activation.J Cell Biol. 2008 Jun 30;181(7):1195-210. doi: 10.1083/jcb.200801078. Epub 2008 Jun 23. J Cell Biol. 2008. PMID: 18573916 Free PMC article.
-
The transcription factor Mef2 links the Drosophila core clock to Fas2, neuronal morphology, and circadian behavior.Neuron. 2013 Jul 24;79(2):281-92. doi: 10.1016/j.neuron.2013.05.015. Neuron. 2013. PMID: 23889933 Free PMC article.
-
p38 and extracellular signal-regulated kinases regulate the myogenic program at multiple steps.Mol Cell Biol. 2000 Jun;20(11):3951-64. doi: 10.1128/MCB.20.11.3951-3964.2000. Mol Cell Biol. 2000. PMID: 10805738 Free PMC article.
-
Transcriptional features of genomic regulatory blocks.Genome Biol. 2009;10(4):R38. doi: 10.1186/gb-2009-10-4-r38. Epub 2009 Apr 19. Genome Biol. 2009. PMID: 19374772 Free PMC article.
References
-
- Black B L, Ligon K L, Zhang Y, Olson E N. Cooperative transcriptional activation by the neurogenic bHLH protein MASH1 and members of the MEF2 family. J Biol Chem. 1996;271:26659–26663. - PubMed
-
- Black B L, Martin J F, Olson E N. The mouse MRF4 promoter is trans-activated directly and indirectly by muscle-specific transcription factors. J Biol Chem. 1995;270:2889–2992. - PubMed
-
- Black, B. L., and E. N. Olson. Unpublished observations.
-
- Bour B A, O’Brien M A, Lockwood W L, Goldstein E S, Bodmer R, Taghert P H, Abmayr S M, Nguyen H T. Drosophila MEF2, a transcription factor that is essential for myogenesis. Genes Dev. 1995;9:730–741. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases