

Myotonic dystrophy kinase-related Cdc42-binding kinase acts as a Cdc42 effector in promoting cytoskeletal reorganization
- PMID: 9418861
- PMCID: PMC121465
- DOI: 10.1128/MCB.18.1.130
Myotonic dystrophy kinase-related Cdc42-binding kinase acts as a Cdc42 effector in promoting cytoskeletal reorganization
Abstract
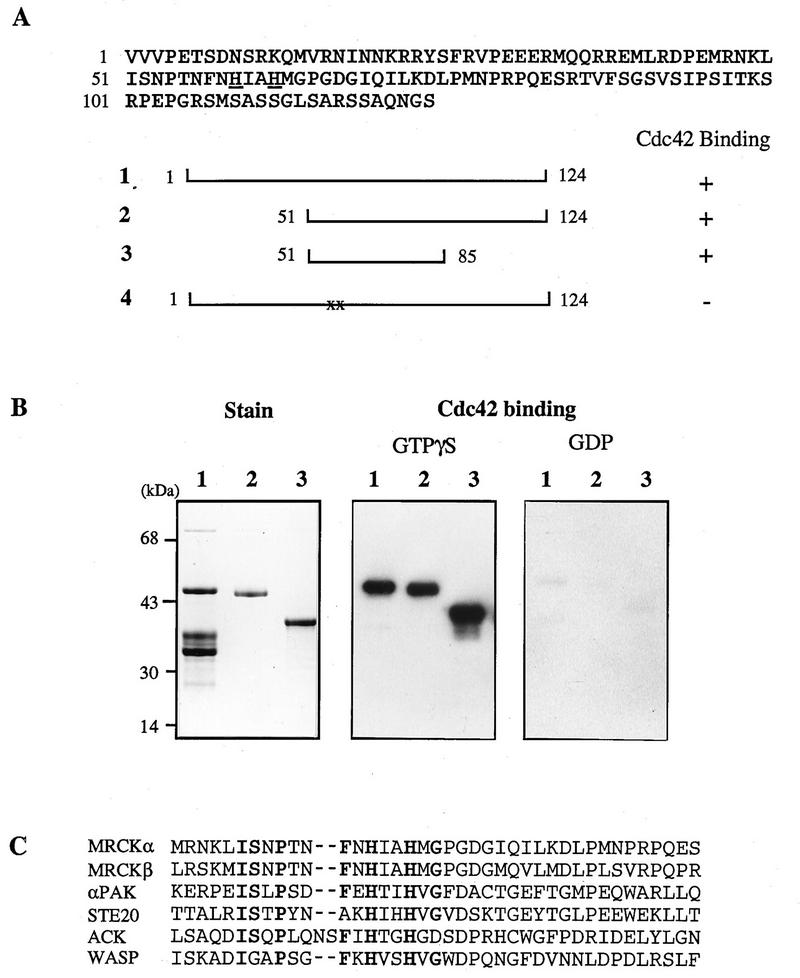
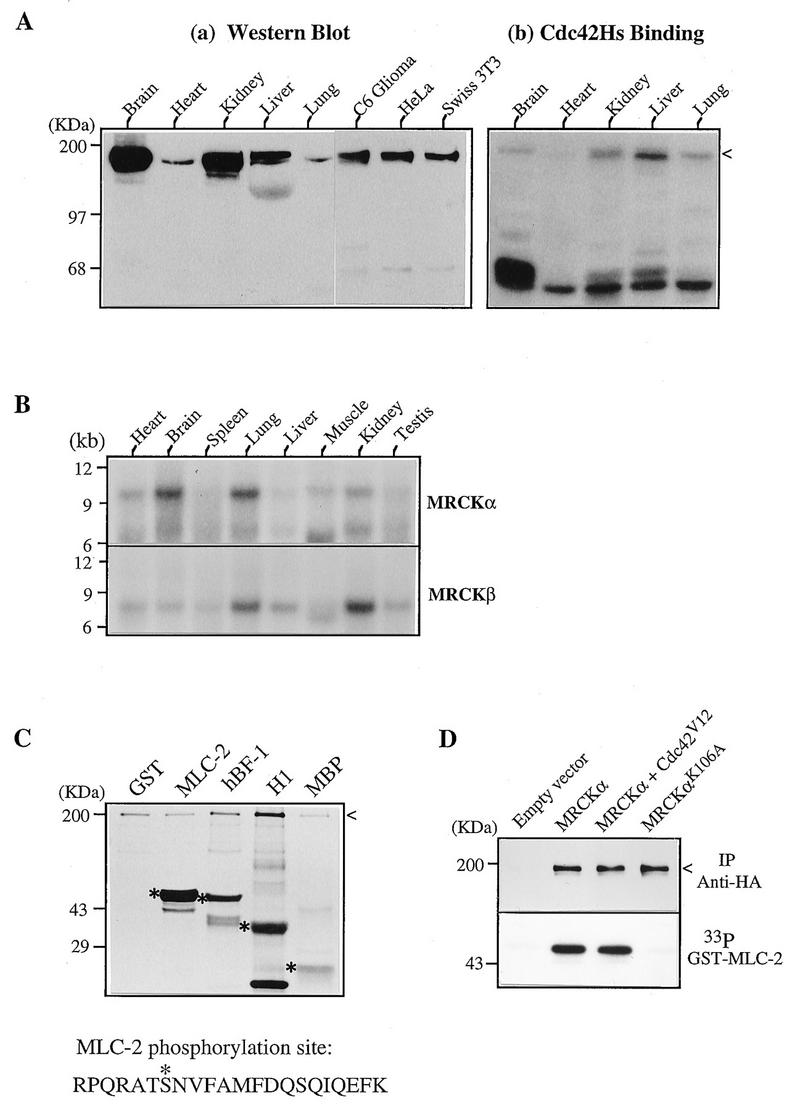
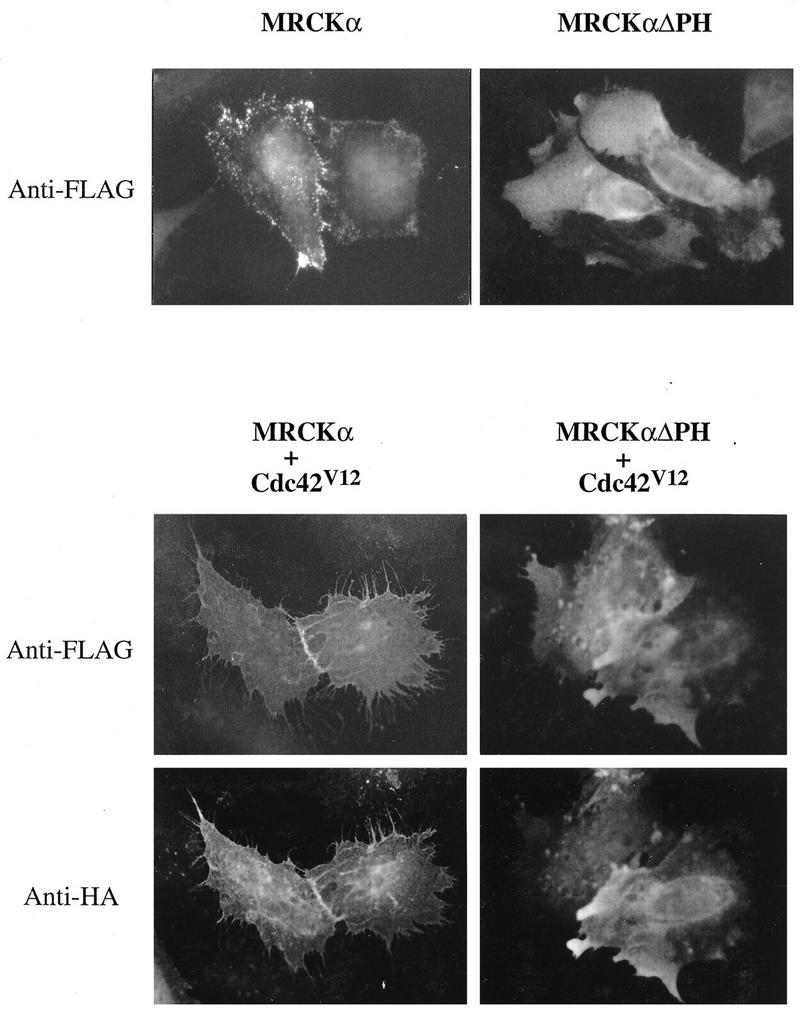
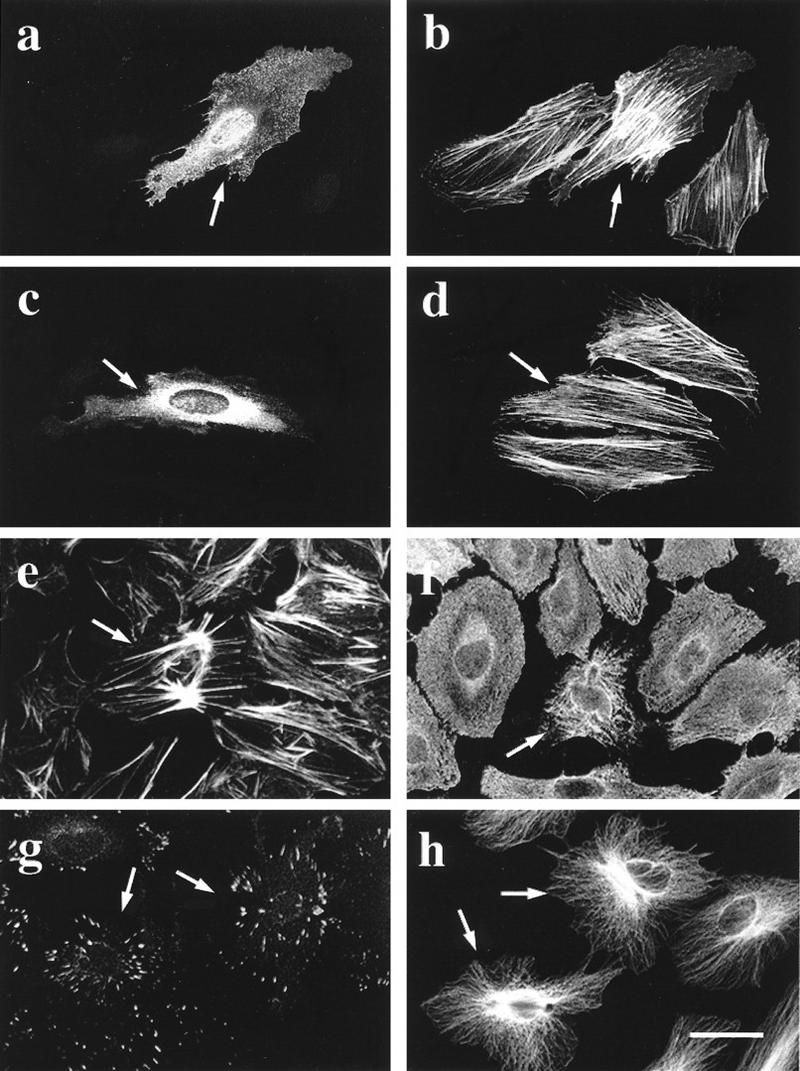
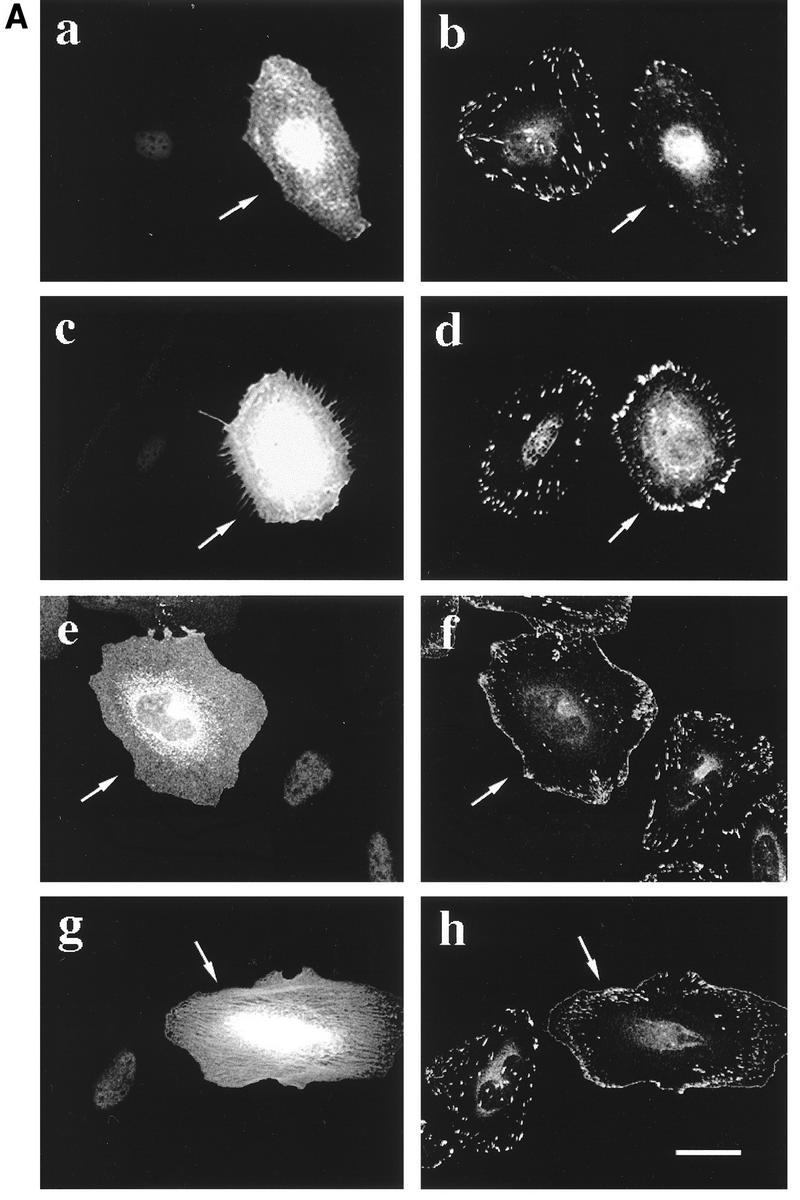
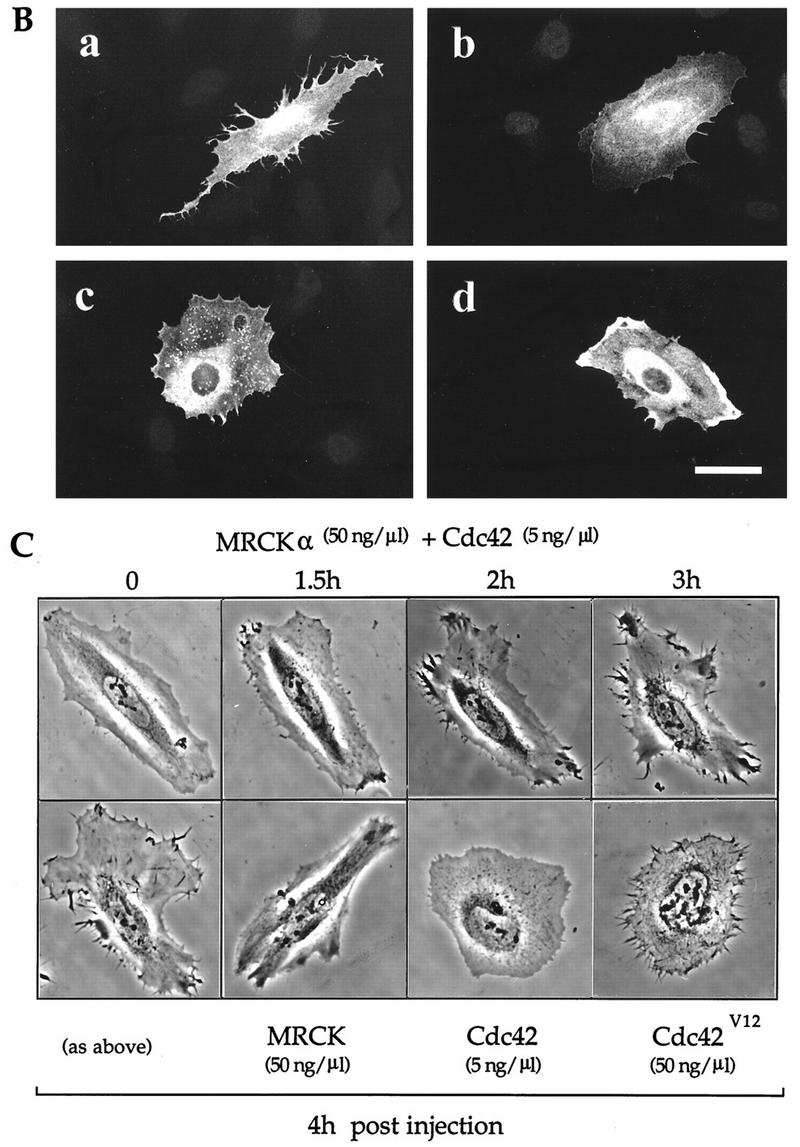
The Rho GTPases play distinctive roles in cytoskeletal reorganization associated with growth and differentiation. The Cdc42/Rac-binding p21-activated kinase (PAK) and Rho-binding kinase (ROK) act as morphological effectors for these GTPases. We have isolated two related novel brain kinases whose p21-binding domains resemble that of PAK whereas the kinase domains resemble that of myotonic dystrophy kinase-related ROK. These approximately 190-kDa myotonic dystrophy kinase-related Cdc42-binding kinases (MRCKs) preferentially phosphorylate nonmuscle myosin light chain at serine 19, which is known to be crucial for activating actin-myosin contractility. The p21-binding domain binds GTP-Cdc42 but not GDP-Cdc42. The multidomain structure includes a cysteine-rich motif resembling those of protein kinase C and n-chimaerin and a putative pleckstrin homology domain. MRCK alpha and Cdc42V12 colocalize, particularly at the cell periphery in transfected HeLa cells. Microinjection of plasmid encoding MRCK alpha resulted in actin and myosin reorganization. Expression of kinase-dead MRCK alpha blocked Cdc42V12-dependent formation of focal complexes and peripheral microspikes. This was not due to possible sequestration of the p21, as a kinase-dead MRCK alpha mutant defective in Cdc42 binding was an equally effective blocker. Coinjection of MRCK alpha plasmid with Cdc42 plasmid, at concentrations where Cdc42 plasmid by itself elicited no effect, led to the formation of the peripheral structures associated with a Cdc42-induced morphological phenotype. These Cdc42-type effects were not promoted upon coinjection with plasmids of kinase-dead or Cdc42-binding-deficient MRCK alpha mutants. These results suggest that MRCK alpha may act as a downstream effector of Cdc42 in cytoskeletal reorganization.
Figures


References
-
- Amano M, Chihara K, Kimura K, Fukata Y, Nakamura N, Matsuura N, Kaibuchi K. Formation of actin stress fibers and focal adhesions enhanced by Rho-kinase. Science. 1997;275:1308–1311. - PubMed
-
- Amano M, Ito M, Kimura K, Fukata Y, Chihara K, Nakano T, Matsuura Y, Kaibuchi K. Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase) J Biol Chem. 1996;271:20246–20249. - PubMed
-
- Amano M, Mukai H, Ono Y, Chihara K, Matsui T, Hamajima Y, Okawa K, Iwamatsu A, Kaibuchi K. Identification of a putative target for Rho as the serine-threonine kinase protein kinase N. Science. 1996;271:648–650. - PubMed
-
- Aspenstrom P, Lindberg U, Hall A. Two GTPases, Cdc42 and Rac, bind directly to a protein implicated in the immunodeficiency disorder Wiskott-Aldrich syndrome. Curr Biol. 1995;6:70–75. - PubMed
-
- Bragodia S, Derijard B, Davis R J, Cerione R A. Cdc42 and PAK-mediated signalling leads to JNK and p38 mitogen-activated protein kinase activation. J Biol Chem. 1995;270:27995–27998. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous