

Mutations of the Drosophila dDP, dE2F, and cyclin E genes reveal distinct roles for the E2F-DP transcription factor and cyclin E during the G1-S transition
- PMID: 9418862
- PMCID: PMC121467
- DOI: 10.1128/MCB.18.1.141
Mutations of the Drosophila dDP, dE2F, and cyclin E genes reveal distinct roles for the E2F-DP transcription factor and cyclin E during the G1-S transition
Erratum in
- Mol Cell Biol 1998 May;18(5):3101
Abstract
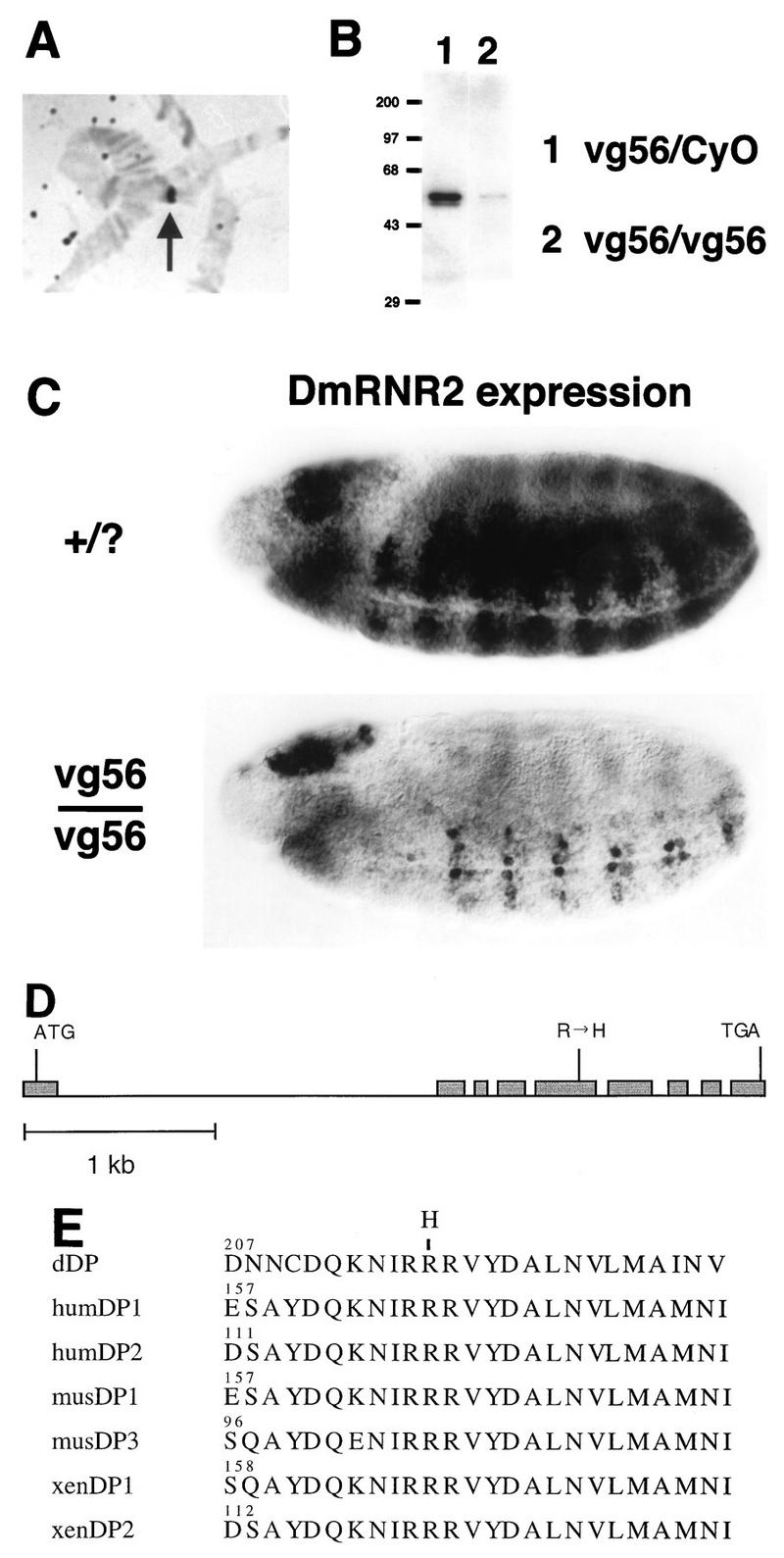
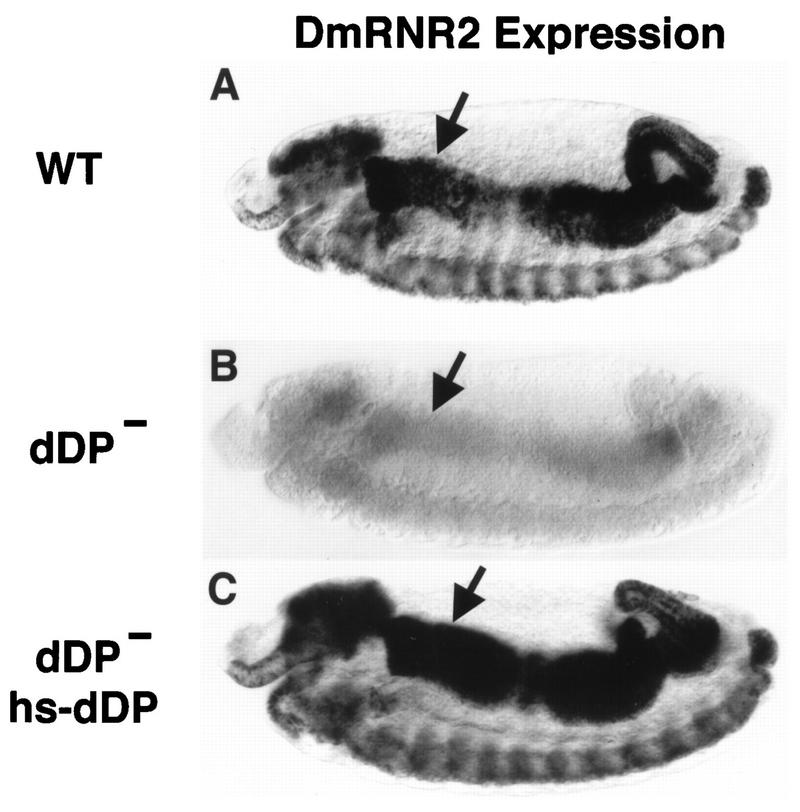
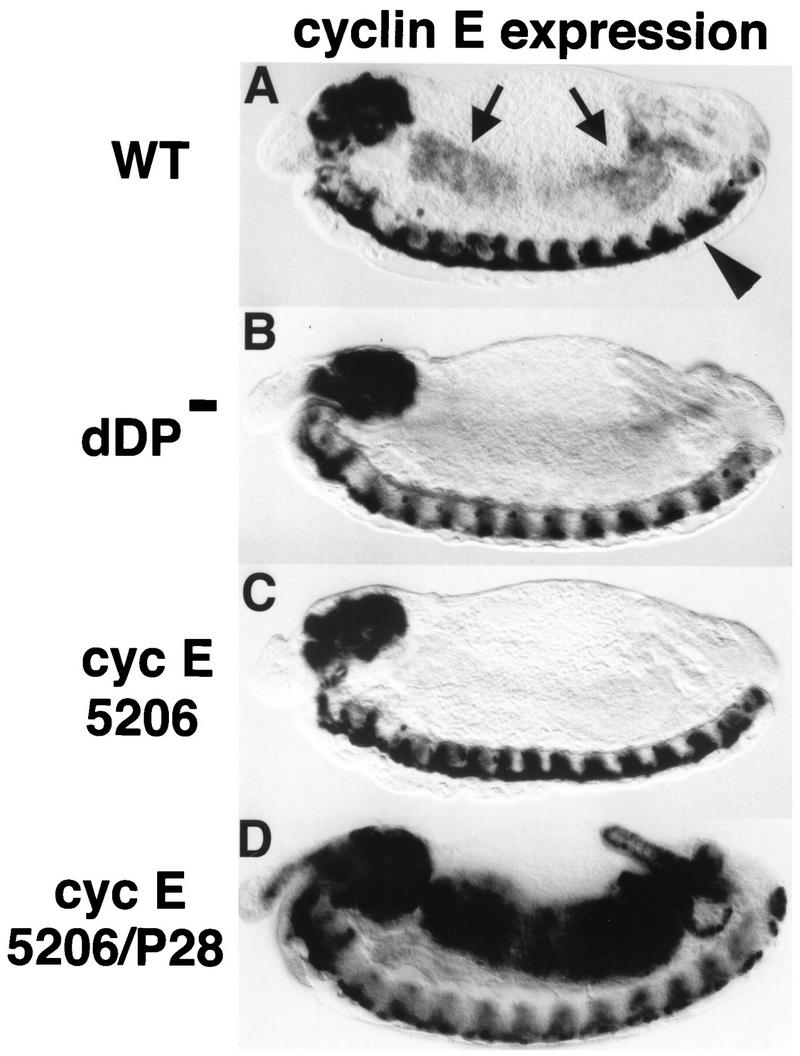
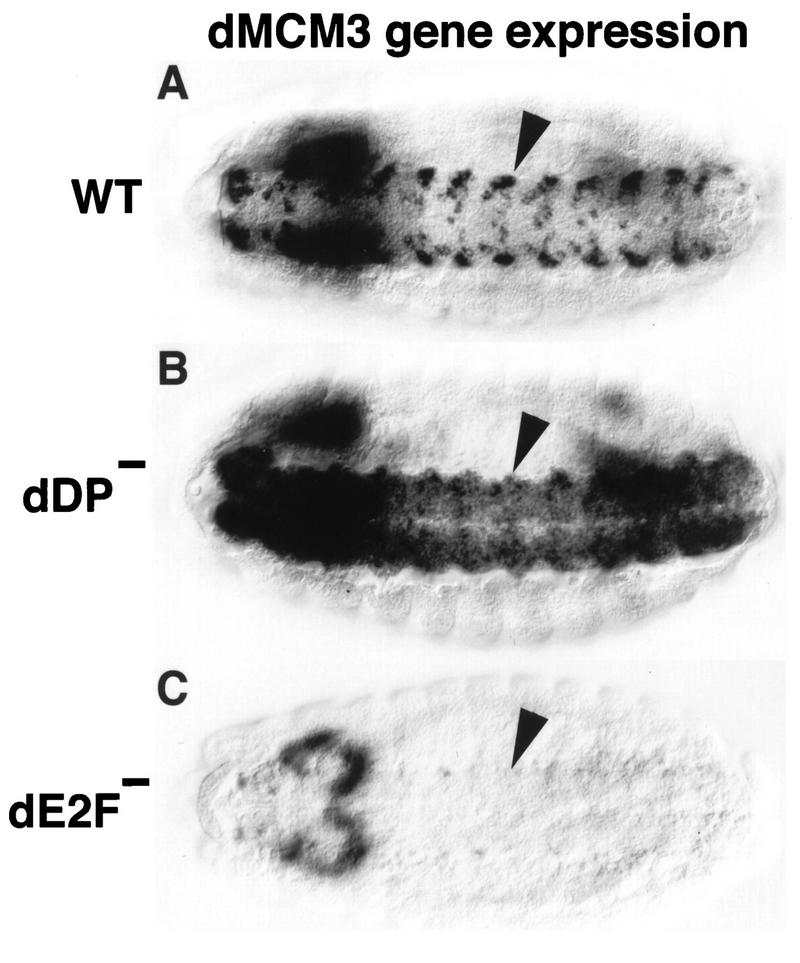
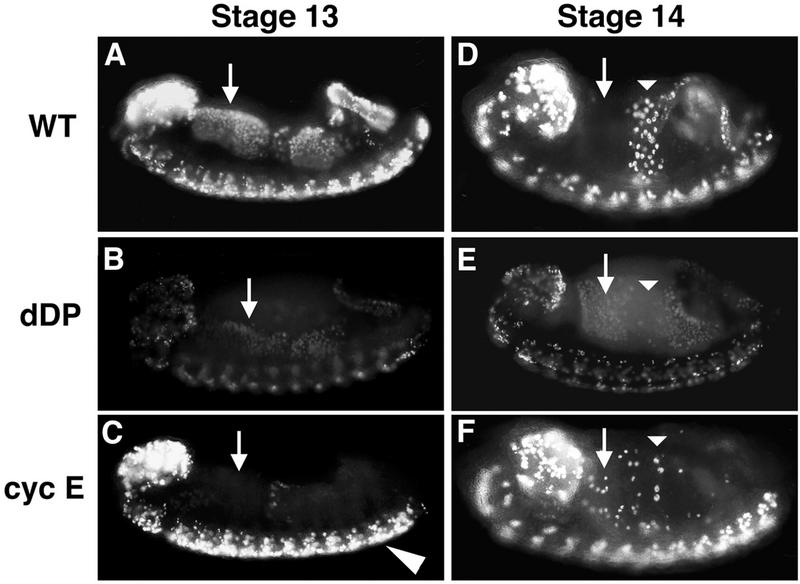
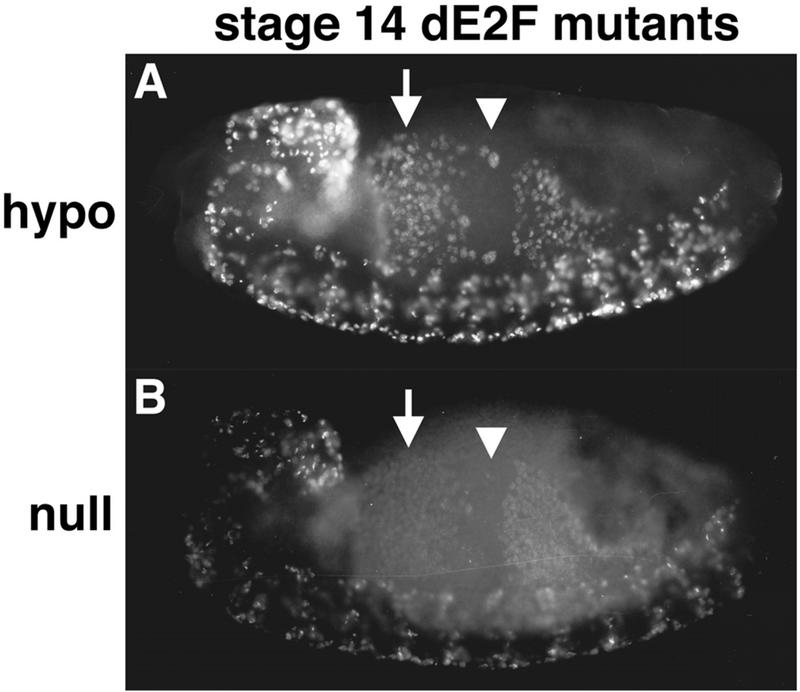
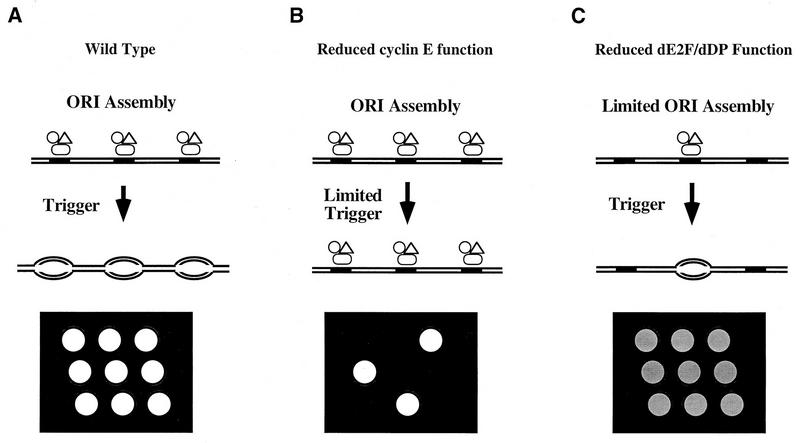
Activation of heterodimeric E2F-DP transcription factors can drive the G1-S transition. Mutation of the Drosophila melanogaster dE2F gene eliminates transcriptional activation of several replication factors at the G1-S transition and compromises DNA replication. Here we describe a mutation in the Drosophila dDP gene. As expected for a defect in the dE2F partner, this mutation blocks G1-S transcription of DmRNR2 and cyclin E as previously described for mutations of dE2F. Mutation of dDP also causes an incomplete block of DNA replication. When S phase is compromised by reducing the activity of dE2F-dDP by either a dE2F or dDP mutation, the first phenotype detected is a reduction in the intensity of BrdU incorporation and a prolongation of the labeling. Notably, in many cells, there was no detected delay in entry into this compromised S phase. In contrast, when cyclin E function was reduced by a hypomorphic allele combination, BrdU incorporation was robust but the timing of S-phase entry was delayed. We suggest that dE2F-dDP contributes to the expression of two classes of gene products: replication factors, whose abundance has a graded effect on replication, and cyclin E, which triggers an all-or-nothing transition from G1 to S phase.
Figures
References
-
- Andrews B J, Herskowitz I. Regulation of cell cycle-dependent gene expression in yeast. J Biol Chem. 1990;265:14057–14060. - PubMed
-
- Ashburner M, Thompson P, Roote J, Lasko P F, Grau Y, El Messal M, Roth S, Simpson P. The genetics of a small autosomal region of Drosophila melanogaster containing the structural gene for alcohol dehydrogenase. VII. Characterization of the region around the snail and cactus loci. Genetics. 1990;126:679–694. - PMC - PubMed
-
- Breeden L. Start-specific transcription in yeast. Curr Top Microbiol Immunol. 1996;208:95–127. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials