

Immunoproteasome assembly: cooperative incorporation of interferon gamma (IFN-gamma)-inducible subunits
- PMID: 9419215
- PMCID: PMC2199179
- DOI: 10.1084/jem.187.1.97
Immunoproteasome assembly: cooperative incorporation of interferon gamma (IFN-gamma)-inducible subunits
Abstract
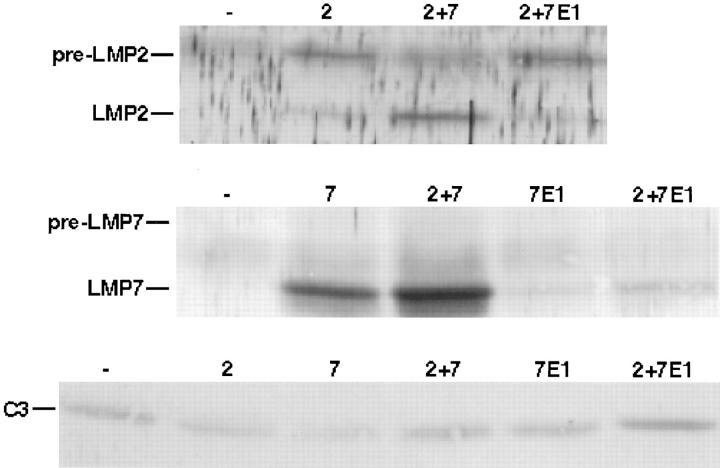
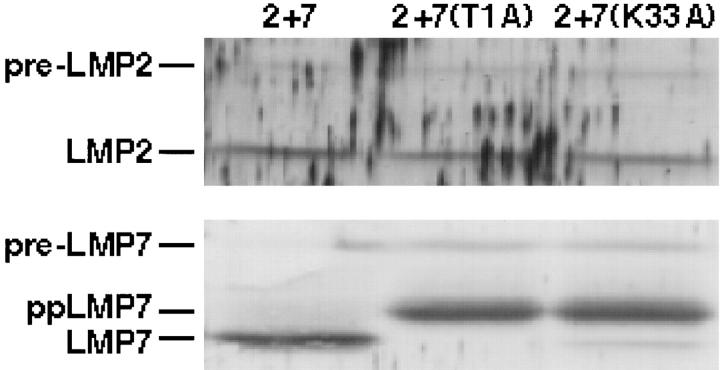
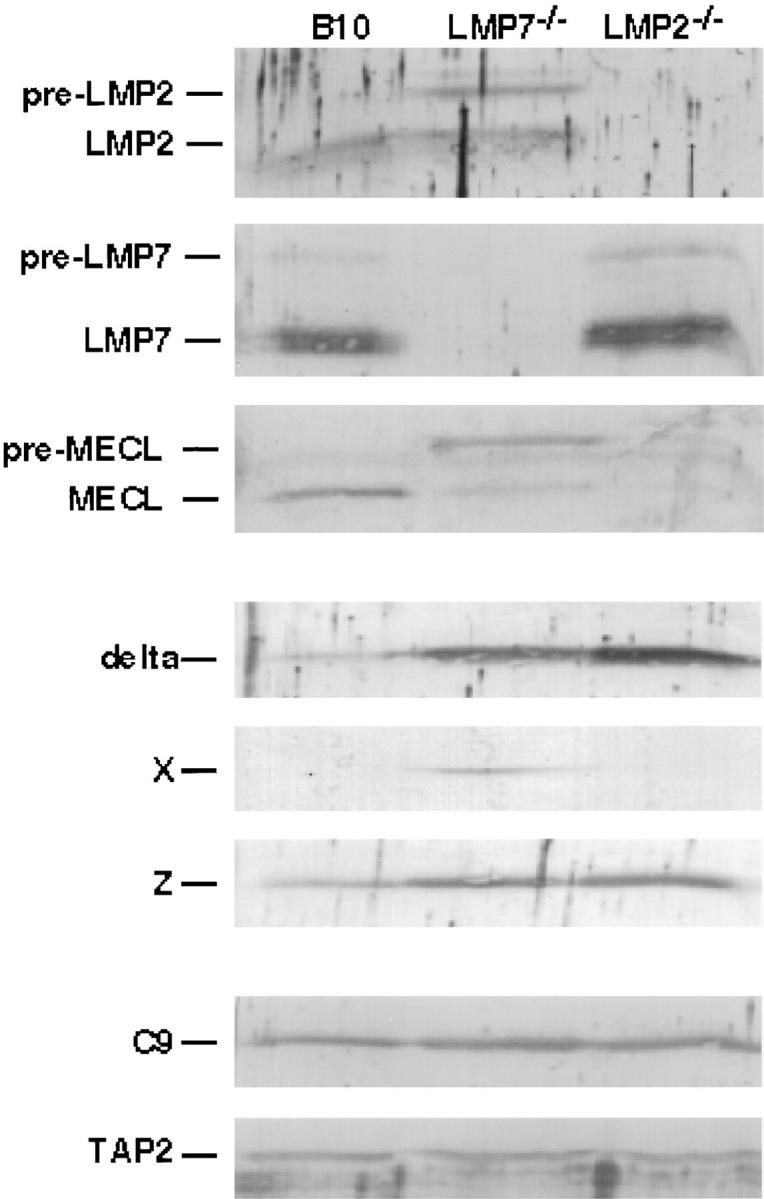
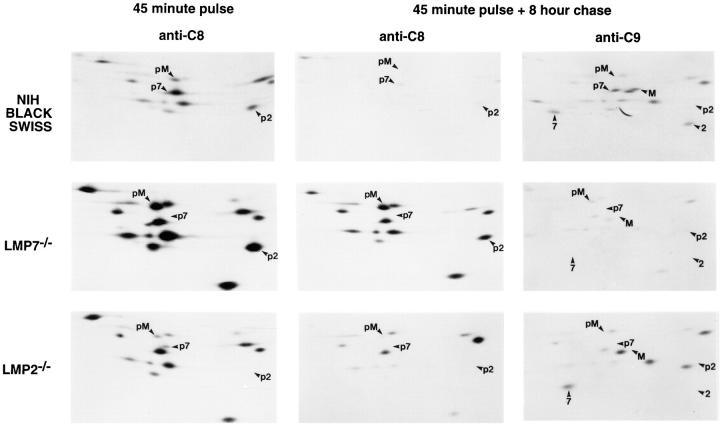
LMP2, LMP7, and MECL are interferon gamma-inducible catalytic subunits of vertebrate 20S proteasomes, which can replace constitutive catalytic subunits (delta, X, and Z, respectively) during proteasome biogenesis. We demonstrate that MECL requires LMP2 for efficient incorporation into preproteasomes, and preproteasomes containing LMP2 and MECL require LMP7 for efficient maturation. The latter effect depends on the presequence of LMP7, but not on LMP7 catalytic activity. This cooperative mechanism favors the assembly of homogeneous "immunoproteasomes" containing all three inducible subunits, suggesting that these subunits act in concert to enhance proteasomal generation of major histocompatibility complex class I-binding peptides.
Figures


References
-
- Coux O, Tanaka K, Goldberg AL. Structure and functions of the 20S and 26S proteasomes. Annu Rev Biochem. 1996;65:801–847. - PubMed
-
- Rock KL, Gramm C, Rothstein L, Clark K, Stein R, Dick L, Hwang D, Goldberg AL. Inhibitors of the proteasome block the degradation of most cell proteins and the generation of peptides presented on MHC class I molecules. Cell. 1994;78:761–771. - PubMed
-
- Groll M, Ditzel L, Löwe J, Stock D, Bochtler M, Bartunik HD, Huber R. Structure of 20S proteasome from yeast at 2.4 Å resolution. Nature. 1997;386:463–471. - PubMed
-
- Seemüller E, Lupas A, Stock D, Löwe J, Huber R, Baumeister W. Proteasome from Thermoplasma acidophilum: a threonine protease. Science. 1995;268:579–582. - PubMed
-
- Martinez CK, Monaco JJ. Homology of proteasome subunits to a major histocompatibility complex–linked LMP gene. Nature. 1991;353:664–667. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
