

Varicella-zoster virus gene 21: transcriptional start site and promoter region
- PMID: 9420198
- PMCID: PMC109347
- DOI: 10.1128/JVI.72.1.42-47.1998
Varicella-zoster virus gene 21: transcriptional start site and promoter region
Abstract
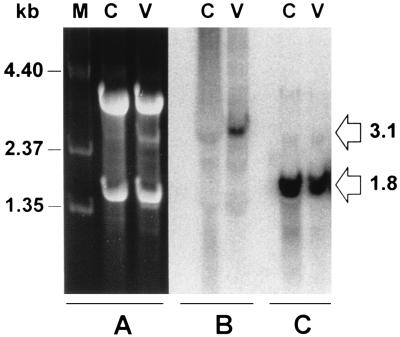
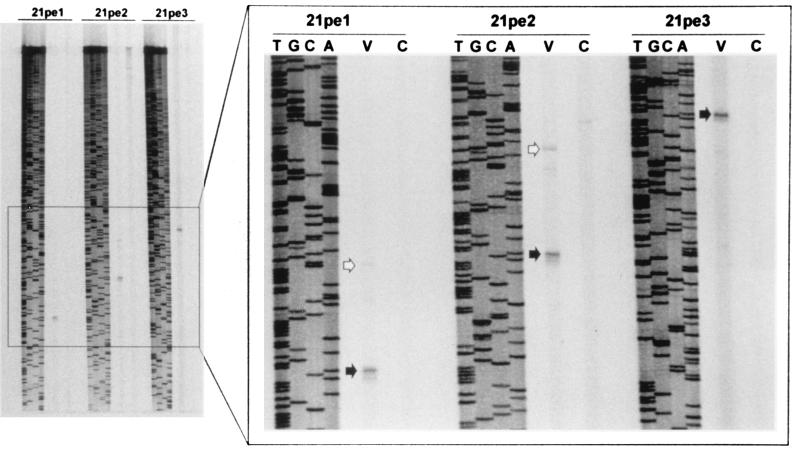
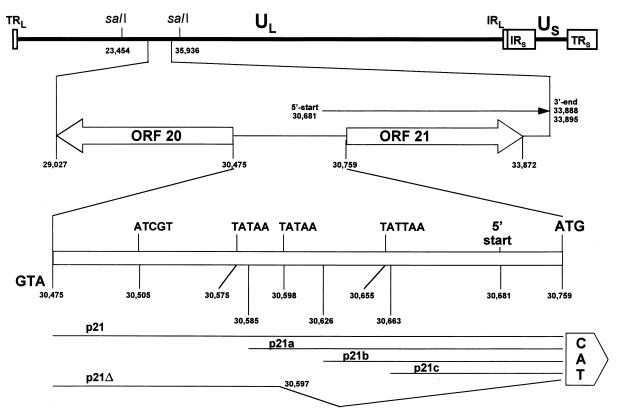
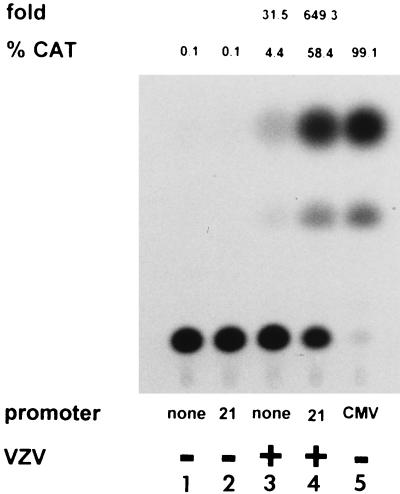
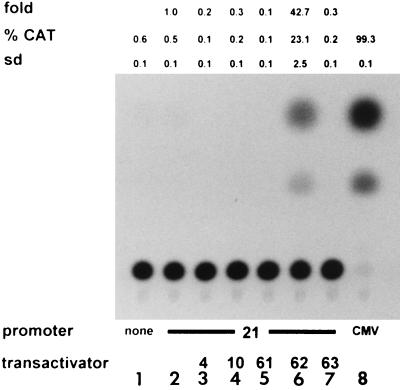
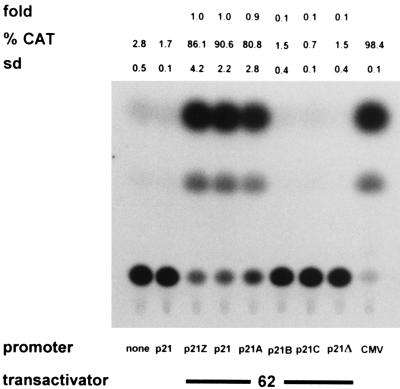
Varicella-zoster virus (VZV) causes chicken pox (varicella), becomes latent in dorsal root ganglia, and reactivates decades later to cause shingles (zoster). During latency, the entire VZV genome is present in a circular form, from which genes 21, 29, 62, and 63 are transcribed. Immediate-early (IE) VZV genes 62 and 63 encode regulators of virus gene transcription, and VZV gene 29 encodes a major DNA-binding protein. However, little is known about the function of VZV gene 21 or the control of its transcription. Using primer extensions, we mapped the start of VZV gene 21 transcription in VZV-infected cells to a single site located at -79 nucleotides (nt) with respect to the initiation codon. To identify the VZV gene 21 promoter, the 284-bp region of VZV DNA separating open reading frames (ORFs) 20 and 21 was cloned upstream from the chloramphenicol acetyltransferase gene. In transient-transfection assays, the VZV gene 21 promoter was transactivated in VZV-infected, but not uninfected, cells. Further, the protein encoded by ORF 62 (IE62), but not those encoded by VZV ORFs 4, 10, 61, and 63, transactivates the VZV gene 21 promoter. By use of transient-cotransfection assays in conjunction with 5' deletions of the VZV gene 21 promoter, a 40-bp segment was shown to be responsible for the transactivation of the VZV gene 21 promoter by IE62. This region was located at -96 to -56 nt with respect to the 5' start of gene 21 transcription.
Figures
Similar articles
-
The varicella-zoster virus (VZV) open-reading frame 29 protein acts as a modulator of a late VZV gene promoter.J Infect Dis. 1998 Nov;178 Suppl 1:S34-8. doi: 10.1086/514276. J Infect Dis. 1998. PMID: 9852970
-
Transcriptional mapping of the varicella-zoster virus regulatory genes encoding open reading frames 4 and 63.J Virol. 1994 Jun;68(6):3570-81. doi: 10.1128/JVI.68.6.3570-3581.1994. J Virol. 1994. PMID: 8189496 Free PMC article.
-
Simian varicella virus gene 28 and 29 promoters share a common upstream stimulatory factor-binding site and are induced by IE62 transactivation.J Gen Virol. 2006 Jun;87(Pt 6):1501-1508. doi: 10.1099/vir.0.81645-0. J Gen Virol. 2006. PMID: 16690914
-
Lessons to be learned from varicella-zoster virus.Vet Microbiol. 1996 Nov;53(1-2):55-66. doi: 10.1016/s0378-1135(96)01234-5. Vet Microbiol. 1996. PMID: 9010998 Review.
-
The role of varicella zoster virus immediate-early proteins in latency and their potential use as components of vaccines.Arch Virol Suppl. 2001;(17):81-9. doi: 10.1007/978-3-7091-6259-0_9. Arch Virol Suppl. 2001. PMID: 11339554 Review.
Cited by
-
In vitro antiviral activity of honey against varicella zoster virus (VZV): A translational medicine study for potential remedy for shingles.Transl Biomed. 2012;3(2):2. doi: 10.3823/434. Transl Biomed. 2012. PMID: 22822475 Free PMC article.
-
Prevalence of varicella-zoster virus DNA in dissociated human trigeminal ganglion neurons and nonneuronal cells.J Virol. 1999 Oct;73(10):8571-7. doi: 10.1128/JVI.73.10.8571-8577.1999. J Virol. 1999. PMID: 10482610 Free PMC article.
-
Promoter activation by the varicella-zoster virus major transactivator IE62 and the cellular transcription factor USF.J Virol. 2006 Aug;80(15):7339-53. doi: 10.1128/JVI.00309-06. J Virol. 2006. PMID: 16840315 Free PMC article.
-
Characterization of varicella-zoster virus gene 21 and 29 proteins in infected cells.J Virol. 2002 Jul;76(14):7228-38. doi: 10.1128/jvi.76.14.7228-7238.2002. J Virol. 2002. PMID: 12072522 Free PMC article.
-
Varicella-zoster virus open reading frame 21, which is expressed during latency, is essential for virus replication but dispensable for establishment of latency.J Virol. 2003 Jan;77(2):1211-8. doi: 10.1128/jvi.77.2.1211-1218.2003. J Virol. 2003. PMID: 12502838 Free PMC article.
References
-
- Ben-Porat T, Rixon F J. Replication of herpesvirus DNA. IV. Analysis of concatemers. Virology. 1979;94:61–70. - PubMed
-
- Betz J L, Wydoski S G. Functional interaction of varicella zoster virus gene 62 protein with the DNA sequence bound by herpes simplex virus ICP4 protein. Virology. 1993;195:793–797. - PubMed
-
- Chirgwin J M, Przybyla A E, MacDonald R J, Rutter W J. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry. 1979;18:5294–5299. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
