

The gH-gL complex of herpes simplex virus (HSV) stimulates neutralizing antibody and protects mice against HSV type 1 challenge
- PMID: 9420201
- PMCID: PMC109350
- DOI: 10.1128/JVI.72.1.65-72.1998
The gH-gL complex of herpes simplex virus (HSV) stimulates neutralizing antibody and protects mice against HSV type 1 challenge
Abstract
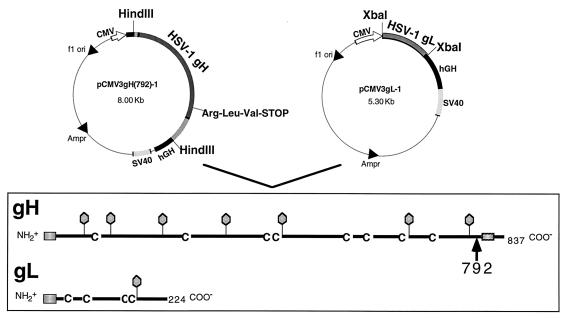
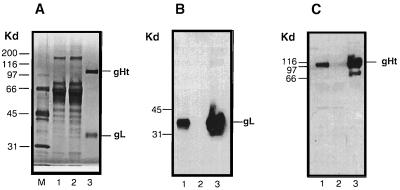
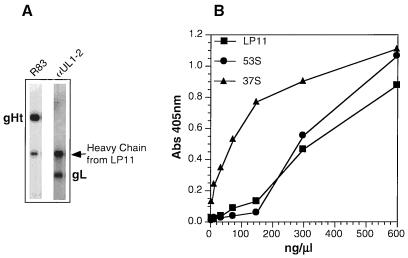
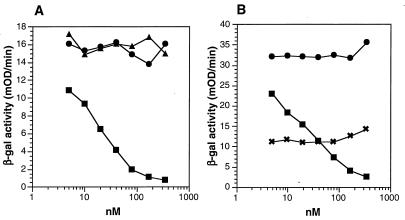
The herpes simplex virus type 1 (HSV-1) gH-gL complex which is found in the virion envelope is essential for virus infectivity and is a major antigen for the host immune system. However, little is known about the precise role of gH-gL in virus entry, and attempts to demonstrate the immunologic or vaccine efficacy of gH and gL separately or as the gH-gL complex have not succeeded. We constructed a recombinant mammalian cell line (HL-7) which secretes a soluble gH-gL complex, consisting of gH truncated at amino acid 792 (gHt) and full-length gL. Purified gHt-gL reacted with gH- and gL-specific monoclonal antibodies, including LP11, which indicates that it retains its proper antigenic structure. Soluble forms of gD (gDt) block HSV infection by interacting with specific cellular receptors. Unlike soluble gD, gHt-gL did not block HSV-1 entry into cells, nor did it enhance the blocking capacity of gD. However, polyclonal antibodies to the complex did block entry even when added after virus attachment. In addition, these antibodies exhibited high titers of complement-independent neutralizing activity against HSV-1. These sera also cross-neutralized HSV-2, albeit at low titers, and cross-reacted with gH-2 present in extracts of HSV-2-infected cells. To test the potential for gHt-gL to function as a vaccine, BALB/c mice were immunized with the complex. As controls, other mice were immunized with gD purified from HSV-infected cells or were sham immunized. Sera from the gD- or gHt-gL-immunized mice exhibited high titers of virus neutralizing activity. Using a zosteriform model of infection, we challenged mice with HSV-1. All animals showed some evidence of infection at the site of virus challenge. Mice immunized with either gD or gHt-gL showed reduced primary lesions and exhibited no secondary zosteriform lesions. The sham-immunized control animals exhibited extensive secondary lesions. Furthermore, mice immunized with either gD or gHt-gL survived virus challenge, while many control animals died. These results suggest that gHt-gL is biologically active and may be a candidate for use as a subunit vaccine.
Figures
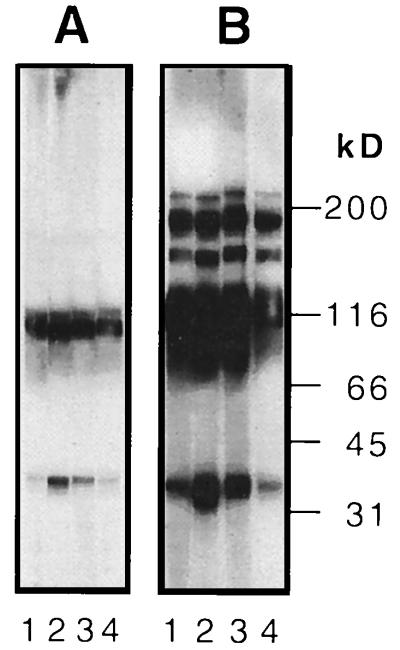
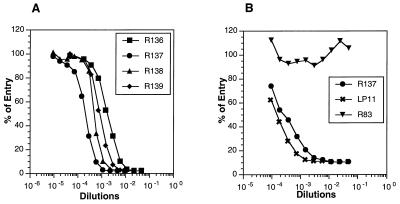
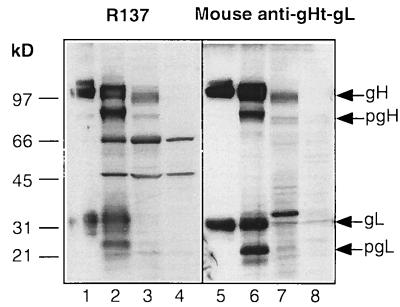
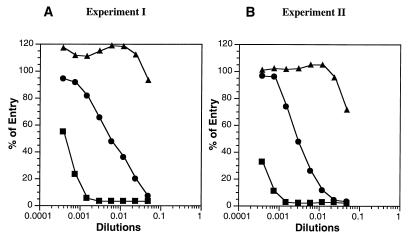
References
-
- Boursnell, M. E. Personal communication.
-
- Browne H, Baxter V, Minson T. Analysis of protective immune responses to the glycoprotein H-glycoprotein L complex of herpes simplex virus type 1. J Gen Virol. 1993;74:2813–2817. - PubMed
-
- Buckmaster E A, Gompels U, Minson A. Characterisation and physical mapping of an HSV-1 glycoprotein of approximately 115 × 103 molecular weight. Virology. 1984;139:408–413. - PubMed
-
- Burke R L. HSV vaccine development. In: Roizman B, Lopez C, editors. The herpesviruses. Vol. 5. New York, N.Y: Raven Press; 1993. pp. 367–380.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
