

Circulating activated platelets reconstitute lymphocyte homing and immunity in L-selectin-deficient mice
- PMID: 9432977
- PMCID: PMC2212105
- DOI: 10.1084/jem.187.2.197
Circulating activated platelets reconstitute lymphocyte homing and immunity in L-selectin-deficient mice
Abstract
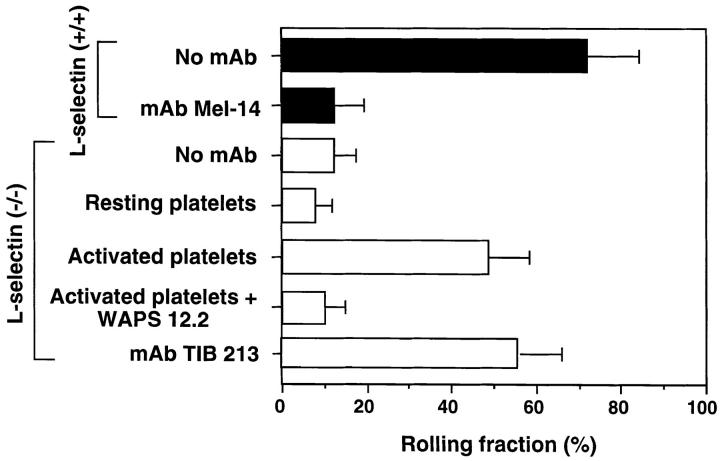
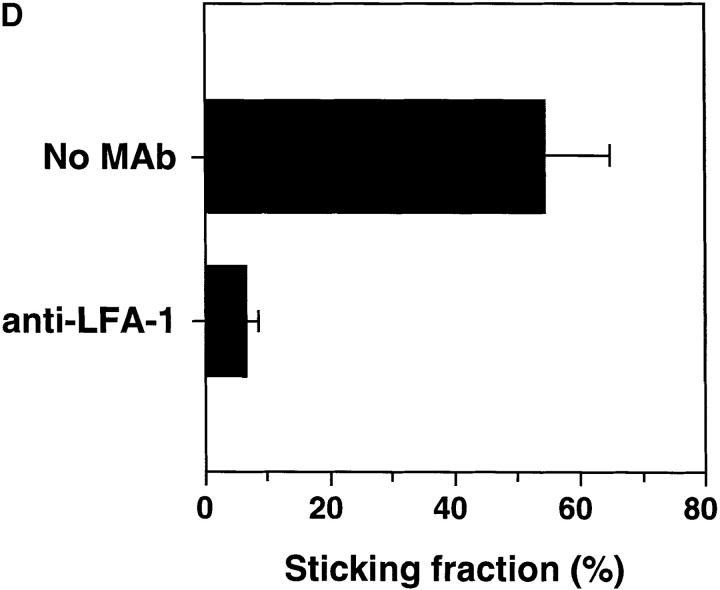
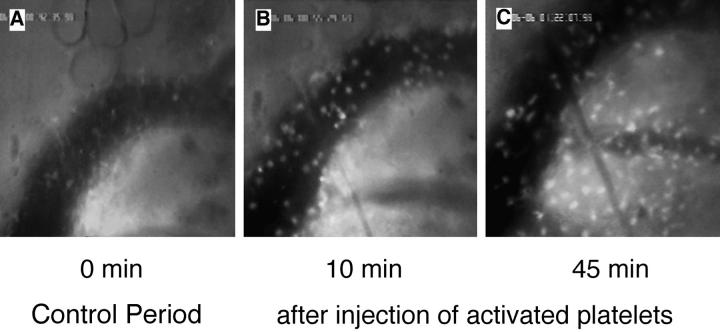
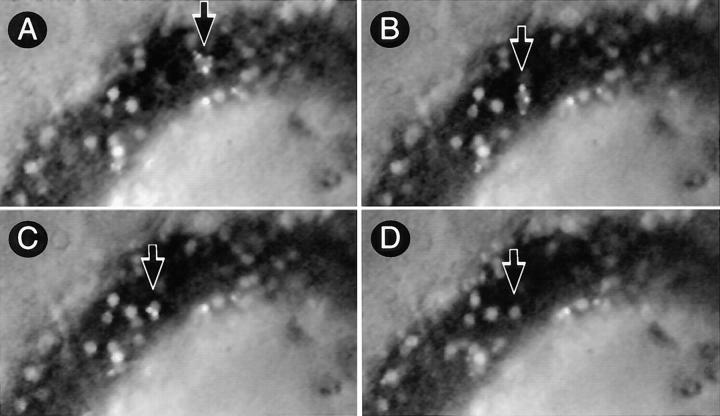
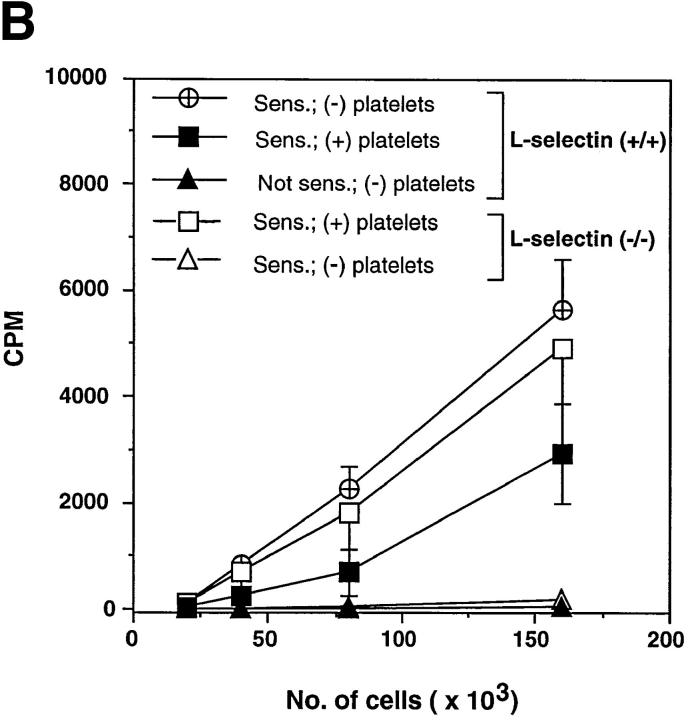
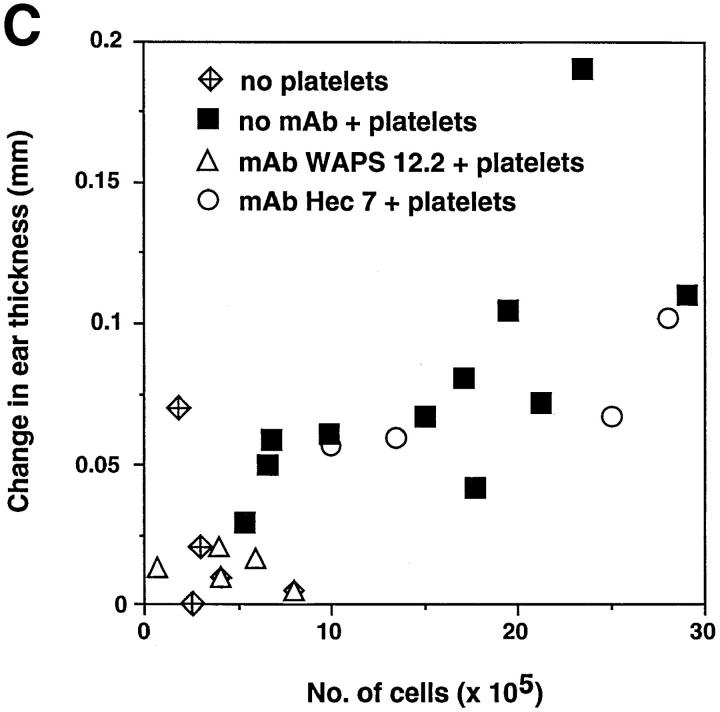
Peripheral lymph nodes (PLN) are critical for immunologic memory formation in response to antigens that penetrate the skin. Blood-borne lymphocytes first encounter such antigens after they home to PLN through a multi-step adhesion process that is normally initiated by L-selectin (CD62L) in high endothelial venules (HEV). Since naive T cells can not enter PLN normally in L-selectin-deficient mice, a delayed type hypersensitivity response to cutaneously applied antigen cannot be mounted. In this study, we report that the administration of activated platelets into the systemic circulation of L-selectin knockout mice restores lymphocyte trafficking to PLN, and reconstitutes T cell-mediated immunity in response to a cutaneous antigen. These effects required platelet-expressed P-selectin that allows activated platelets to transiently form a bridge between lymphocytes and HEV, thereby enabling lymphocytes to undergo subsequent beta2 integrin-dependent firm adhesion. These profound effects of platelet-mediated cell-cell interactions on lymphocyte trafficking and formation of immunologic memory may impact on a variety of autoimmune and inflammatory conditions.
Figures
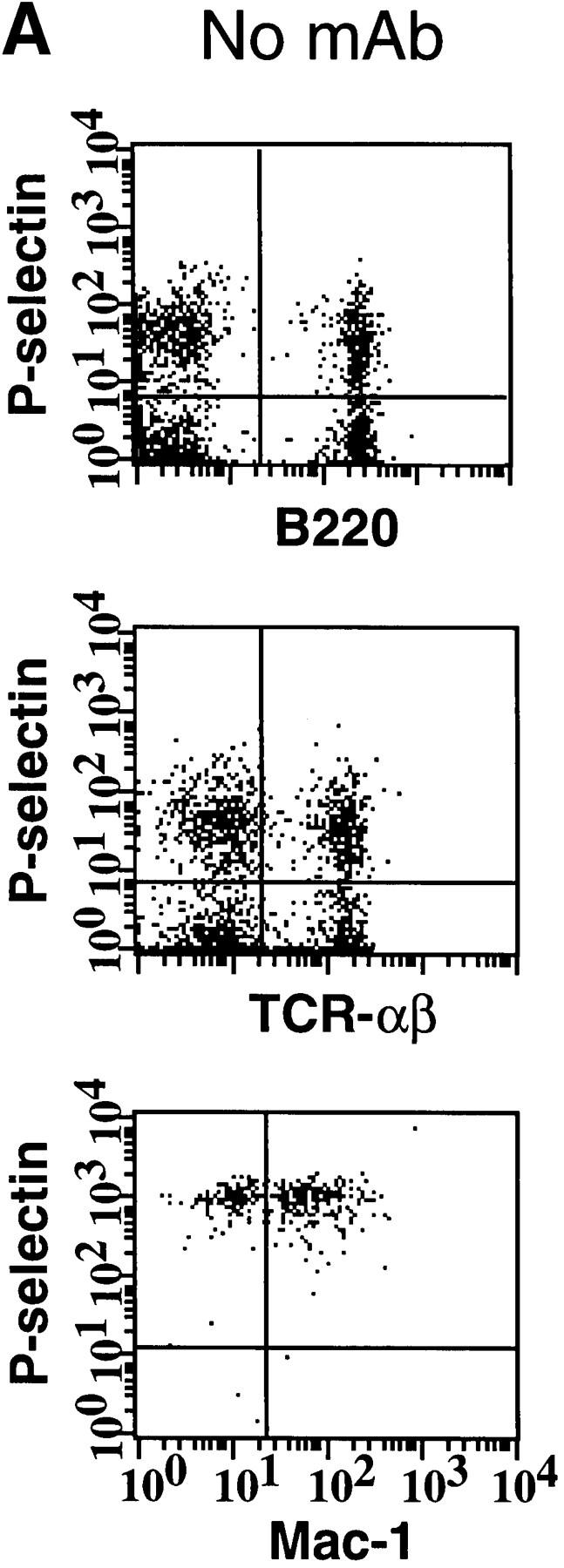
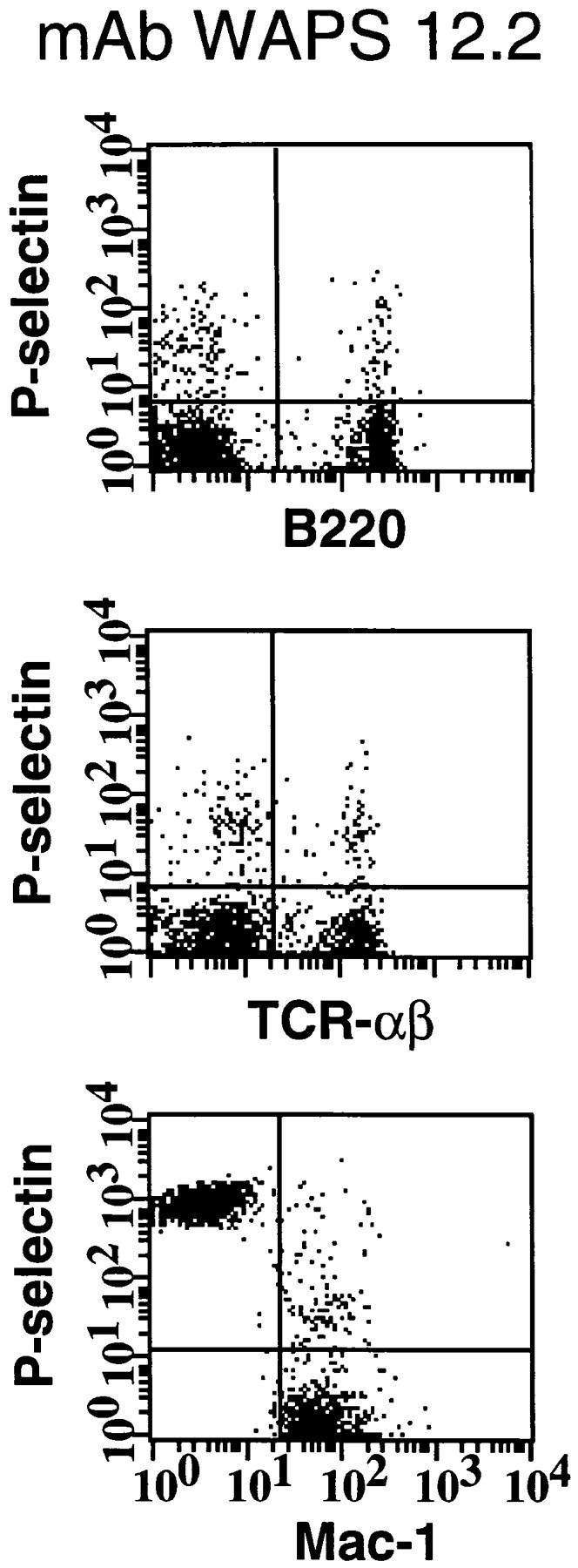
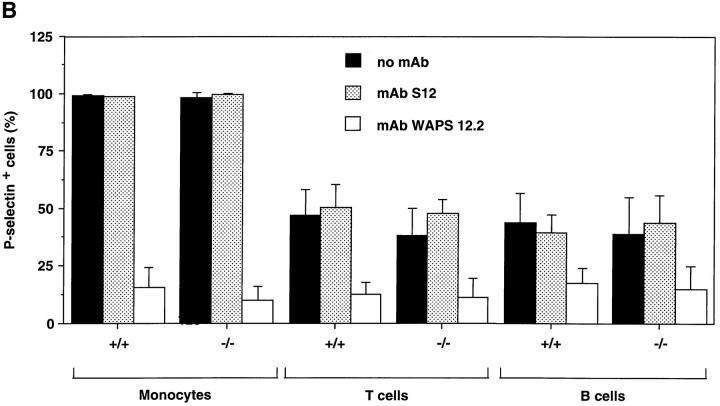
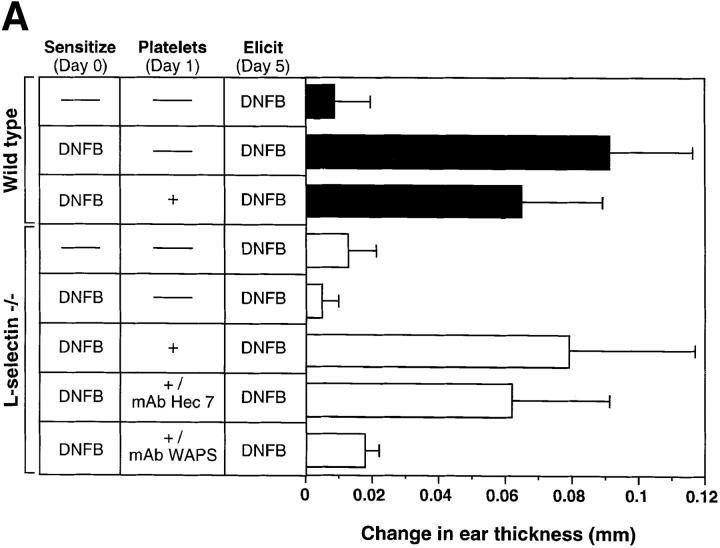
Similar articles
-
Lyphocyte migration in L-selectin-deficient mice. Altered subset migration and aging of the immune system.J Immunol. 1996 Aug 1;157(3):1096-106. J Immunol. 1996. PMID: 8757614
-
Molecular mechanisms of lymphocyte homing to peripheral lymph nodes.J Exp Med. 1998 Jan 19;187(2):205-16. doi: 10.1084/jem.187.2.205. J Exp Med. 1998. PMID: 9432978 Free PMC article.
-
In situ analysis of lymphocyte migration to lymph nodes.Cell Adhes Commun. 1998;6(2-3):85-96. doi: 10.3109/15419069809004463. Cell Adhes Commun. 1998. PMID: 9823458 Review.
-
Platelet-mediated lymphocyte delivery to high endothelial venules.Science. 1996 Jul 12;273(5272):252-5. doi: 10.1126/science.273.5272.252. Science. 1996. PMID: 8662511
-
L-selectin: role in regulating homeostasis and cutaneous inflammation.J Dermatol Sci. 2009 Dec;56(3):141-7. doi: 10.1016/j.jdermsci.2009.10.001. Epub 2009 Nov 3. J Dermatol Sci. 2009. PMID: 19889515 Free PMC article. Review.
Cited by
-
Adaptive immune responses mediated by natural killer cells.Immunol Rev. 2010 May;235(1):286-96. doi: 10.1111/j.0105-2896.2010.00906.x. Immunol Rev. 2010. PMID: 20536570 Free PMC article. Review.
-
P-Selectin: Basic Aspects.J Thromb Thrombolysis. 1998 Sep;6(2):93-102. doi: 10.1023/A:1008837218655. J Thromb Thrombolysis. 1998. PMID: 10751790 No abstract available.
-
Delayed mortality and attenuated thrombocytopenia associated with severe malaria in urokinase- and urokinase receptor-deficient mice.Infect Immun. 2000 Jul;68(7):3822-9. doi: 10.1128/IAI.68.7.3822-3829.2000. Infect Immun. 2000. PMID: 10858190 Free PMC article.
-
Understanding high endothelial venules: Lessons for cancer immunology.Oncoimmunology. 2015 May 7;4(6):e1008791. doi: 10.1080/2162402X.2015.1008791. eCollection 2015 Jun. Oncoimmunology. 2015. PMID: 26155419 Free PMC article. Review.
-
The role of platelets in the pathogenesis of systemic sclerosis.Front Immunol. 2012 Jun 18;3:160. doi: 10.3389/fimmu.2012.00160. eCollection 2012. Front Immunol. 2012. PMID: 22719739 Free PMC article.
References
-
- Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. - PubMed
-
- Gallatin WM, Weissman IL, Butcher EC. A cell-surface molecule involved in organ-specific homing of lymphocytes. Nature. 1983;304:30–34. - PubMed
-
- von Andrian UH. Intravital microscopy of the peripheral lymph node microcirculation in mice. Microcirculation. 1996;3:287–300. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
