

Tissue-type plasminogen activator is a target of the tumor suppressor gene maspin
- PMID: 9435220
- PMCID: PMC18448
- DOI: 10.1073/pnas.95.2.499
Tissue-type plasminogen activator is a target of the tumor suppressor gene maspin
Abstract
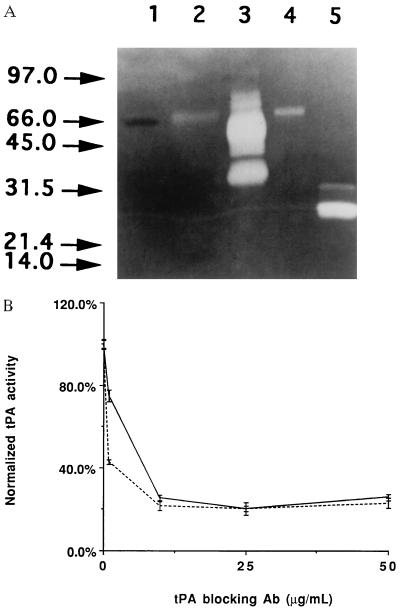
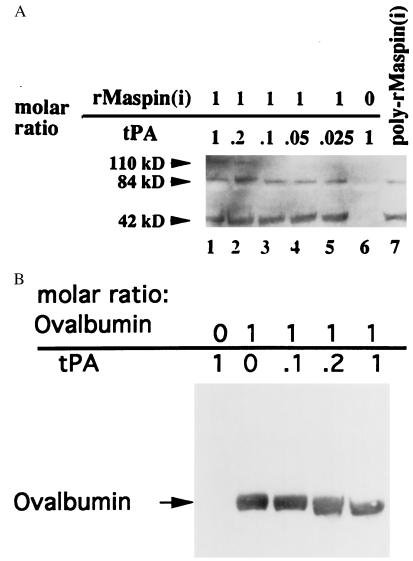
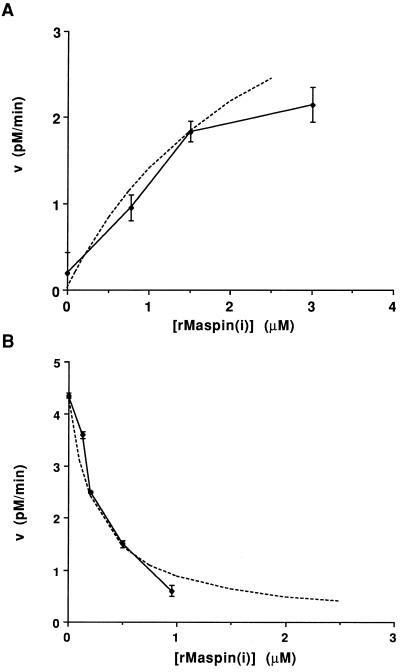
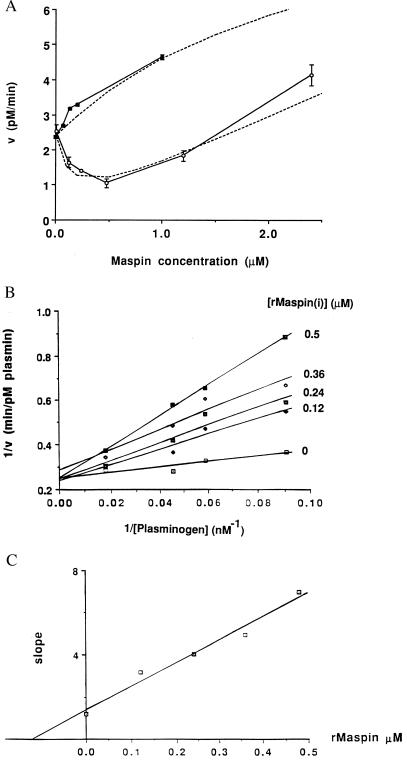
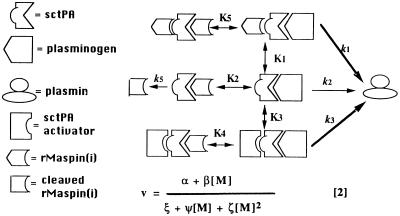
The maspin protein has tumor suppressor activity in breast and prostate cancers. It inhibits cell motility and invasion in vitro and tumor growth and metastasis in nude mice. Maspin is structurally a member of the serpin (serine protease inhibitors) superfamily but deviates somewhat from classical serpins. We find that single-chain tissue plasminogen activator (sctPA) specifically interacts with the maspin reactive site loop peptide and forms a stable complex with recombinant maspin [rMaspin(i)]. Major effects of rMaspin(i) are observed on plasminogen activation by sctPA. First, rMaspin(i) activates free sctPA. Second, it inhibits sctPA preactivated by poly-D-lysine. Third, rMaspin(i) exerts a biphasic effect on the activity of sctPA preactivated by fibrinogen/gelatin, acting as a competitive inhibitor at low concentrations (< 0.5 microM) and as a stimulator at higher concentrations. Fourth, 38-kDa C-terminal truncated rMaspin(i) further stimulates fibrinogen/gelatin-associated sctPA. rMaspin(i) acts specifically; it does not inhibit urokinase-type plasminogen activator, plasmin, chymotrypsin, trypsin, or elastase. Our kinetic data are quantitatively consistent with a model in which two segregated domains of maspin interact with the catalytic and activating domains of sctPA. These complex interactions between maspin and sctPA in vitro suggest a mechanism by which maspin regulates plasminogen activation by sctPA bound to the epithelial cell surface.
Figures
Similar articles
-
The surface of prostate carcinoma DU145 cells mediates the inhibition of urokinase-type plasminogen activator by maspin.Cancer Res. 2000 Sep 1;60(17):4771-8. Cancer Res. 2000. PMID: 10987285
-
Pleiotrophic inhibition of pericellular urokinase-type plasminogen activator system by endogenous tumor suppressive maspin.Cancer Res. 2001 Dec 15;61(24):8676-82. Cancer Res. 2001. PMID: 11751384
-
Maspin inhibits cell migration in the absence of protease inhibitory activity.J Biol Chem. 2002 Dec 6;277(49):46845-8. doi: 10.1074/jbc.C200532200. Epub 2002 Oct 15. J Biol Chem. 2002. PMID: 12384513
-
A role of novel serpin maspin in tumor progression: the divergence revealed through efforts to converge.J Cell Physiol. 2006 Dec;209(3):631-5. doi: 10.1002/jcp.20786. J Cell Physiol. 2006. PMID: 17001674 Review.
-
Maspin--a novel protease inhibitor with tumor-suppressing activity in breast cancer.Acta Oncol. 2000;39(8):931-4. doi: 10.1080/02841860050215909. Acta Oncol. 2000. PMID: 11206999 Review.
Cited by
-
Expression of maspin is associated with the intestinal type of gastric adenocarcinoma.Cancer Res Treat. 2005 Aug;37(4):228-32. doi: 10.4143/crt.2005.37.4.228. Epub 2005 Aug 31. Cancer Res Treat. 2005. PMID: 19956519 Free PMC article.
-
Maspin and MCM2 immunoprofiling in salivary gland carcinomas.Diagn Pathol. 2011 Sep 26;6:89. doi: 10.1186/1746-1596-6-89. Diagn Pathol. 2011. PMID: 21943228 Free PMC article.
-
Pathobiological significance of vascular endothelial growth factor and Maspin expressions in human gastric carcinoma.World J Gastroenterol. 2004 Sep 15;10(18):2624-7. doi: 10.3748/wjg.v10.i18.2624. World J Gastroenterol. 2004. PMID: 15309707 Free PMC article.
-
Elevated maspin expression is associated with better overall survival in esophageal squamous cell carcinoma (ESCC).PLoS One. 2013 May 22;8(5):e63581. doi: 10.1371/journal.pone.0063581. Print 2013. PLoS One. 2013. PMID: 23717449 Free PMC article.
-
Identification of an intrinsic determinant critical for maspin subcellular localization and function.PLoS One. 2013 Nov 21;8(11):e74502. doi: 10.1371/journal.pone.0074502. eCollection 2013. PLoS One. 2013. PMID: 24278104 Free PMC article.
References
-
- Zou Z, Anisowics A, Hendrix M J C, Thor A, Neveu M, Sheng S, Rafidi K, Seftor E A, Sager R. Science. 1994;263:526–529. - PubMed
-
- Sheng S, Pemberton P A, Sager R. J Biol Chem. 1994;269:30988–30993. - PubMed
-
- Sager R, Sheng S, Pemberton P, Hendrix M J C. Curr Top Microbiol Immunol. 1996;213:51–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
