

Chromatin assembly factor I contributes to the maintenance, but not the re-establishment, of silencing at the yeast silent mating loci
- PMID: 9436982
- PMCID: PMC316446
- DOI: 10.1101/gad.12.2.219
Chromatin assembly factor I contributes to the maintenance, but not the re-establishment, of silencing at the yeast silent mating loci
Abstract
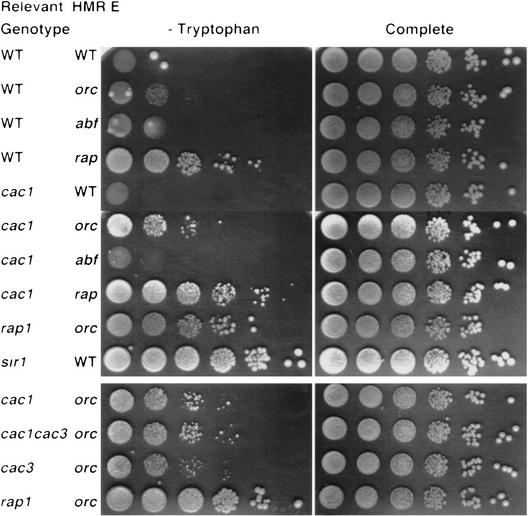
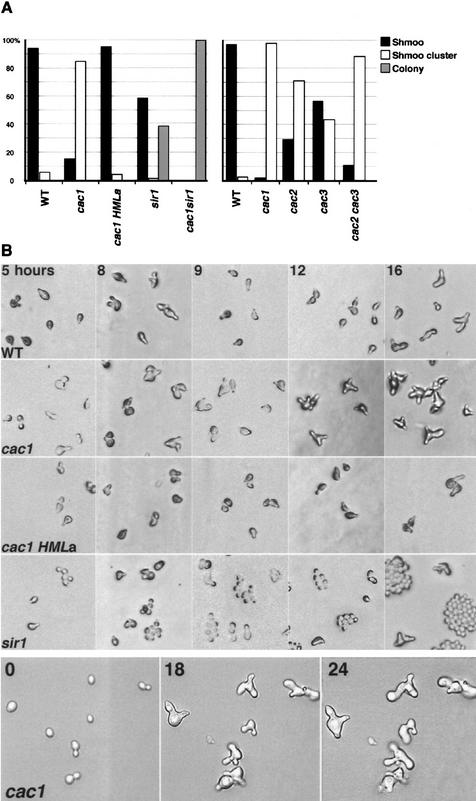
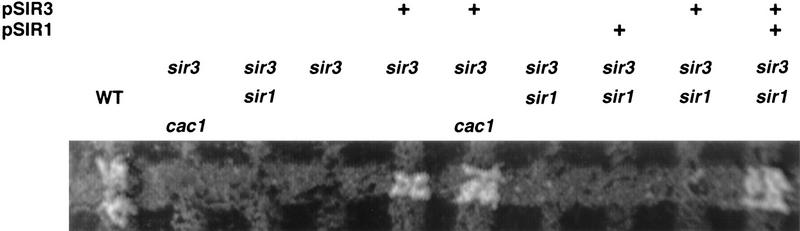
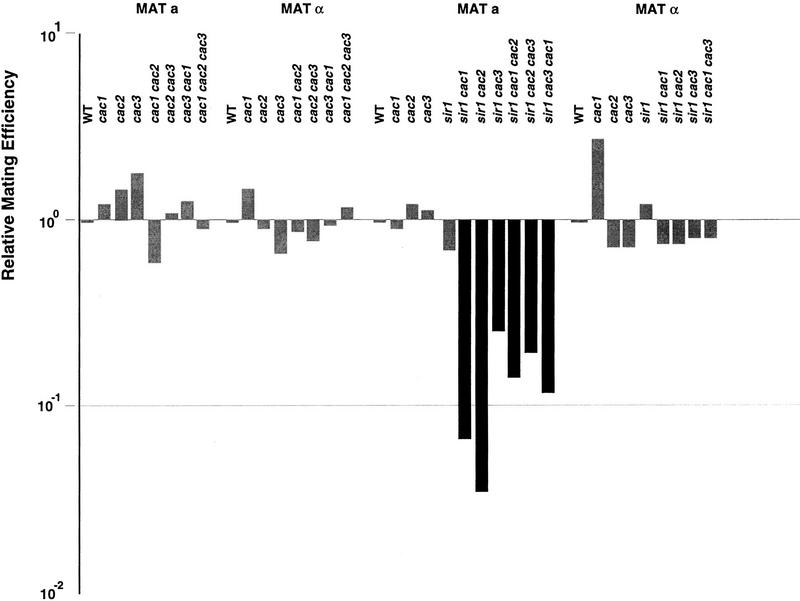
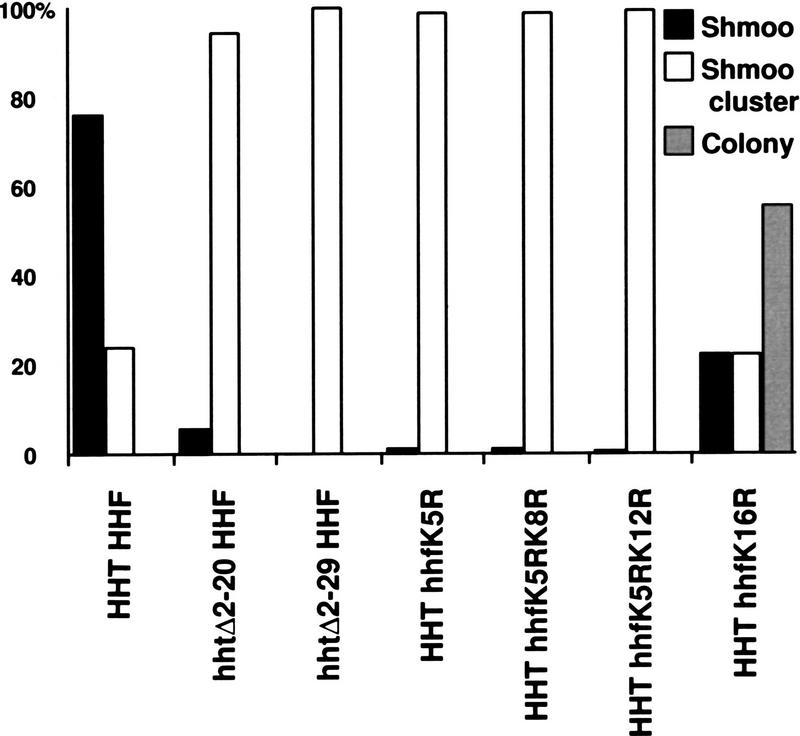
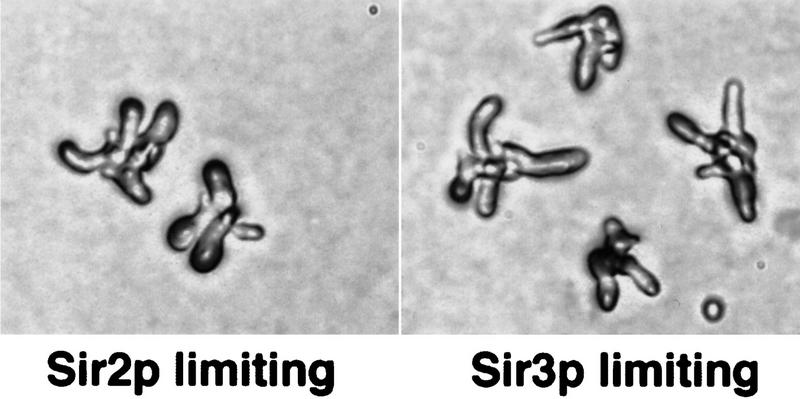
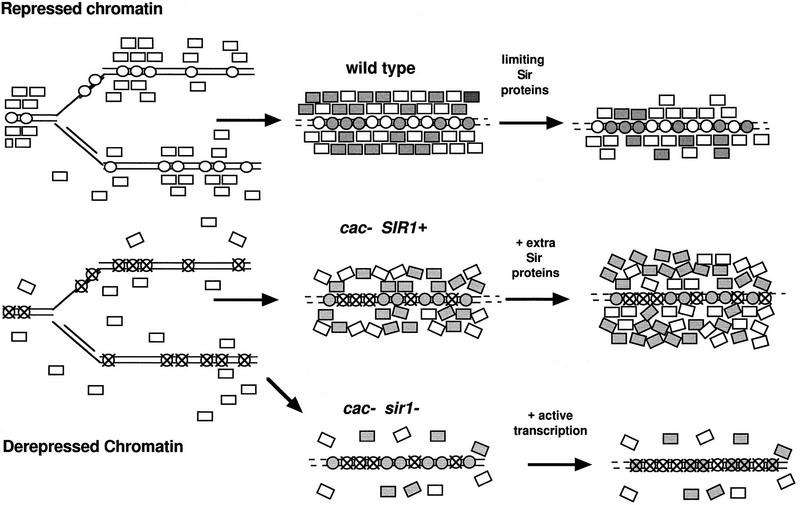
CAC1/RLF2 encodes the largest subunit of chromatin assembly factor I (CAF-I), a complex that assembles newly synthesized histones onto recently replicated DNA in vitro. In vivo, cac1/rlf2 mutants are defective in telomeric silencing and mislocalize Rap1p, a telomere-binding protein. Here, we report that in cells lacking CAF-I the silent mating loci are derepressed partially. MATa cac1 cells exhibit an unusual response to alpha-factor: They arrest and form mating projections (shmoos) initially, but are unable to sustain the arrest state, giving rise to clusters of shmooing cells. cac1 MATa HMLa HMRa strains do not form these shmoo clusters, indicating that derepression of HMLalpha causes the shmoo cluster phenotype in cac1 cells. When SIR3 is reintroduced into sir1 sir3 cells, HML remains derepressed indicating that SIR1 is required for the re-establishment of silencing at HML. In contrast, when SIR3 is reintroduced into cac1 sir3 cells, silencing is restored to HML, indicating that CAF-I is not required for the re-establishment of silencing. Loss of the other CAF-I subunits (Cac2p and Cac3p/Msi1p) also results in the shmoo cluster phenotype, implying that loss of CAF-I activity gives rise to this unstable repression of HML. Strains carrying certain mutations in the amino terminus of histone H4 and strains with limiting amounts of Sir2p or Sir3p also form shmoo clusters, implying that the shmoo cluster phenotype is indicative of defects in maintenance of the structural integrity of silent chromatin. MATa cac- sir1 double mutants have a synergistic mating defect, suggesting that the two silencing mechanisms, establishment and maintenance, function cooperatively. We propose a model to explain the distinctions between the establishment and the maintenance of silent chromatin.
Figures
References
-
- Bell SP, Kobayashi R, Stillman B. Yeast origin recognition complex functions in transcription silencing and DNA replication. Science. 1993;262:1844–1849. - PubMed
-
- Brand AH, Micklem G, Nasmyth K. A yeast silencer contains sequences that can promote autonomous plasmid replication and transcriptional activation. Cell. 1987;51:709–719. - PubMed
-
- Buck SW, Shore D. Action of a RAP1 carboxy-terminal silencing domain reveals an underlying competition between HMR and telomeres in yeast. Genes & Dev. 1995;9:370–384. - PubMed
-
- Chien C-t, Buck S, Sternglanz R, Shore D. Targeting of SIR1 protein establishes transcriptional silencing at HM loci and telomeres in yeast. Cell. 1993;75:531–555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases