

Regulation of POU genes by castor and hunchback establishes layered compartments in the Drosophila CNS
- PMID: 9436984
- PMCID: PMC316437
- DOI: 10.1101/gad.12.2.246
Regulation of POU genes by castor and hunchback establishes layered compartments in the Drosophila CNS
Abstract
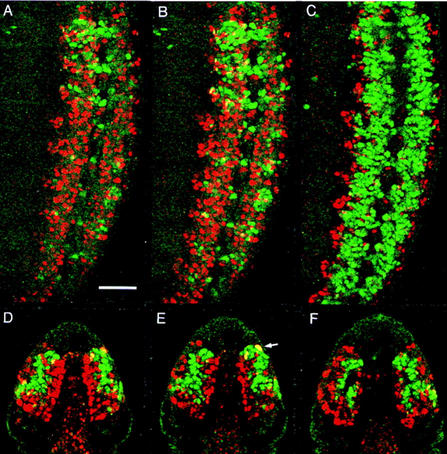
POU transcription factors participate in cell-identity decisions during nervous system development, yet little is known about the regulatory networks controlling their expression. We report all known Drosophila POU genes require castor (cas) for correct CNS expression. drifter and I-POU depend on cas for full expression, whereas pdm-1 and pdm-2 are negatively regulated. cas encodes a zinc finger protein that shares DNA-binding specificity with another pdm repressor: the gap segmentation gene regulator Hunchback (Hb). Our studies reveal that the embryonic CNS contains sequentially generated neuroblast sublineages that can be distinguished by their expression of either Hb, Pdm-1, or Cas. Hb and Cas may directly silence pdm expression in early and late developing sublineages, given that pdm-1 cis-regulatory DNA contains >=32 Hb/Cas-binding sites and its enhancer(s) are ectopically activated in cas- neuroblasts. In addition, the targeted misexpression of Cas in all neuroblast lineages reduces Pdm-1 expression without altering Hb expression. By ensuring correct POU gene expression boundaries, hb and cas maintain temporal subdivisions in the cell-identity circuitry controlling CNS development.
Figures








References
-
- Anderson MG, Perkins GL, Chittick P, Shrigley RJ, Johnson WA. drifter, a Drosophila POU-domain transcription factor, is required for correct differentiation and migration of tracheal cells and midline glia. Genes & Dev. 1995;9:123–137. - PubMed
-
- Artavanis-Tsakonas SA, Matsuno K, Fortini ME. Notch Signaling. Science. 1995;268:225–232. - PubMed
-
- Ashburner M. Drosophila: A laboratory manual. Cold Sping Harbor, NY: Cold Spring Harbor Laboratory Press; 1989.
-
- Bhat K, Schedl P. The Drosophila miti-mere gene, a member of the POU family, is required for the specification of the RP2/sibling lineage during neurogenesis. Development. 1994;120:1483–1501. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous