

Plasma membrane localization of G alpha z requires two signals
- PMID: 9436987
- PMCID: PMC25209
- DOI: 10.1091/mbc.9.1.1
Plasma membrane localization of G alpha z requires two signals
Abstract
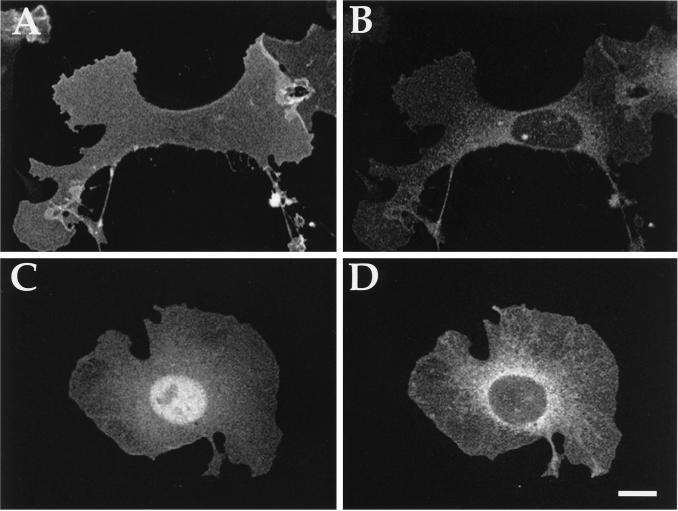
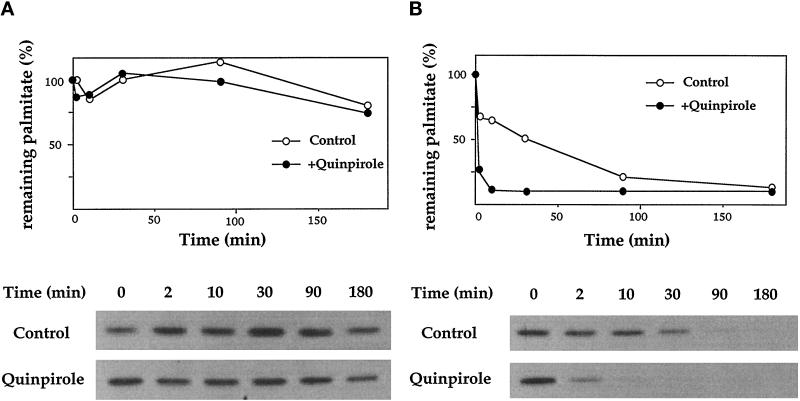
Three covalent attachments anchor heterotrimeric G proteins to cellular membranes: the alpha subunits are myristoylated and/or palmitoylated, whereas the gamma chain is prenylated. Despite the essential role of these modifications in membrane attachment, it is not clear how they cooperate to specify G protein localization at the plasma membrane, where the G protein relays signals from cell surface receptors to intracellular effector molecules. To explore this question, we studied the effects of mutations that prevent myristoylation and/or palmitoylation of an epitope-labeled alpha subunit, alpha z. Wild-type alpha z (alpha z-WT) localizes specifically at the plasma membrane. A mutant that incorporates only myristate is mistargeted to intracellular membranes, in addition to the plasma membrane, but transduces hormonal signals as well as does alpha z-WT. Removal of the myristoylation site produced a mutant alpha z that is located in the cytosol, is not efficiently palmitoylated, and does not relay the hormonal signal. Coexpression of beta gamma with this myristoylation defective mutant transfers it to the plasma membrane, promotes its palmitoylation, and enables it to transmit hormonal signals. Pulse-chase experiments show that the palmitate attached to this myristoylation-defective mutant turns over much more rapidly than does palmitate on alpha z-WT, and that the rate of turnover is further accelerated by receptor activation. In contrast, receptor activation does not increase the slow rate of palmitate turnover on alpha z-WT. Together these results suggest that myristate and beta gamma promote stable association with membranes not only by providing hydrophobicity, but also by stabilizing attachment of palmitate. Moreover, palmitoylation confers on alpha z specific localization at the plasma membrane.
Figures



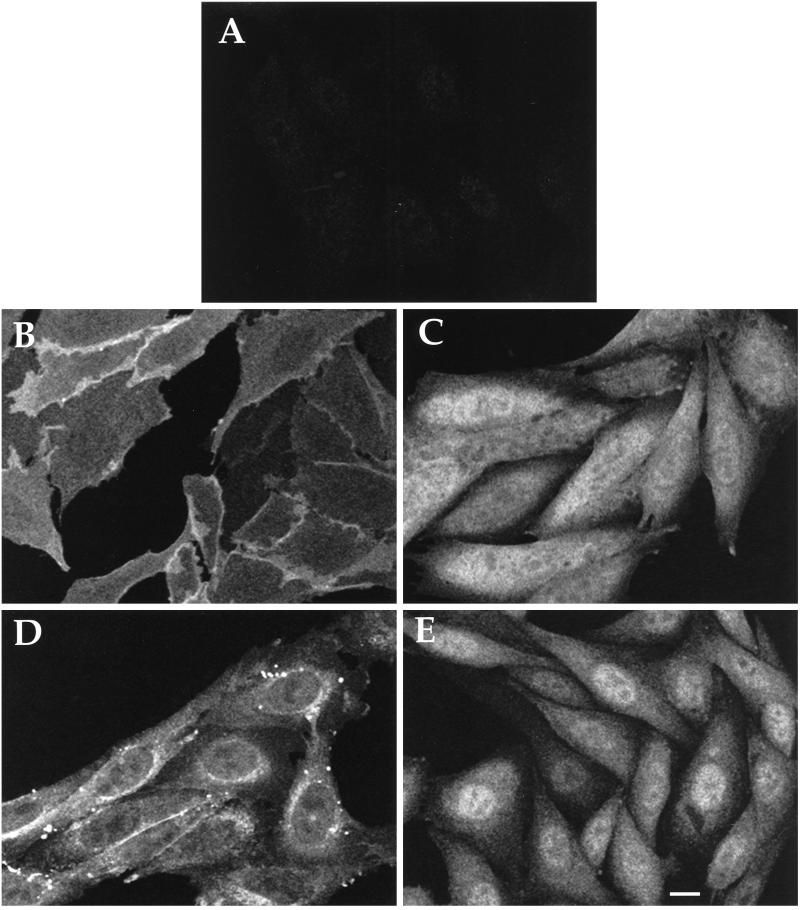

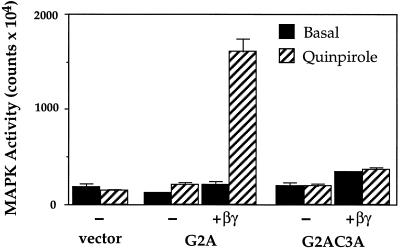
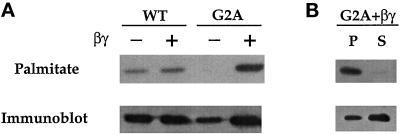
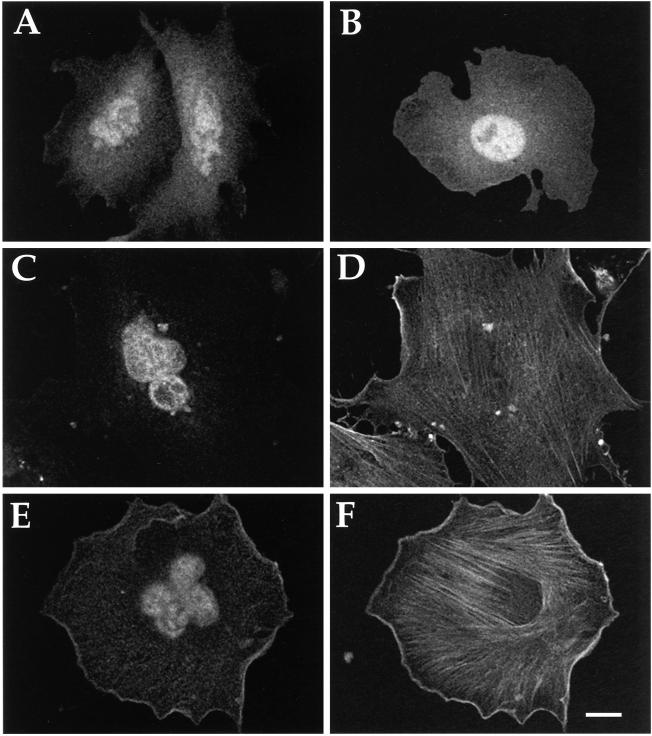
References
-
- Berthiaume L, Resh MD. Biochemical characterization of a palmitoyl acyltransferase activity that palmitoylates myristoylated proteins. J Biol Chem. 1995;270:22399–22405. - PubMed
-
- Bhatnagar RS, Gordon JI. Understanding covalent modifications of proteins by lipids: where cell biology and biophysics mingle. Trends Cell Biol. 1997;7:14–20. - PubMed
-
- Bigay J, Faurobert E, Franco M, Chabre M. Roles of lipid modifications of transducin subunits in their GDP-dependent association and membrane binding. Biochemistry. 1994;33:14081–14090. - PubMed
-
- Boman AL, Kahn RA. Arf proteins: the membrane traffic police? Trends Biochem Sci. 1995;20:147–150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
