

A fungal kinesin required for organelle motility, hyphal growth, and morphogenesis
- PMID: 9436993
- PMCID: PMC25223
- DOI: 10.1091/mbc.9.1.89
A fungal kinesin required for organelle motility, hyphal growth, and morphogenesis
Abstract
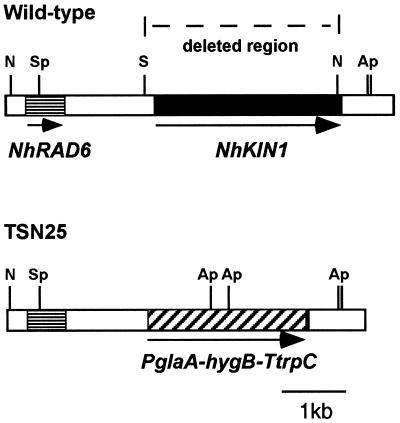
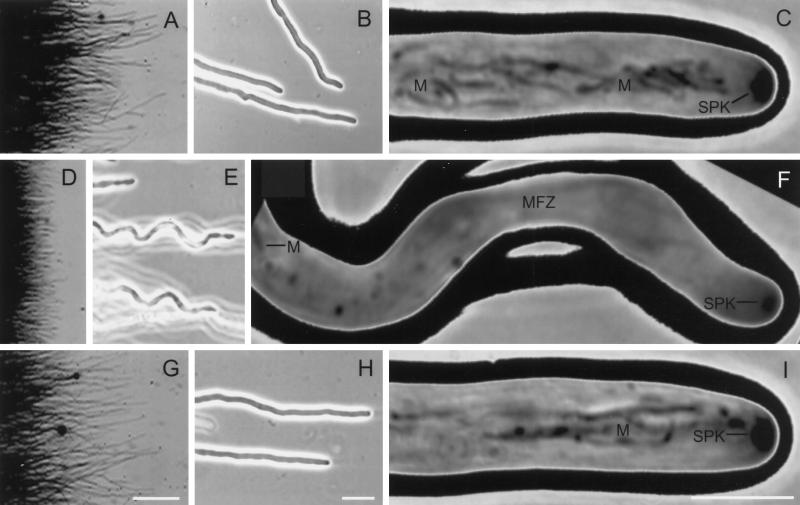
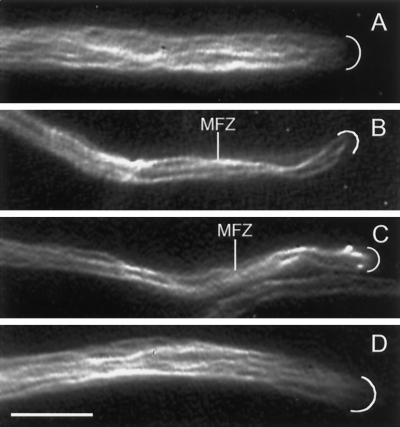
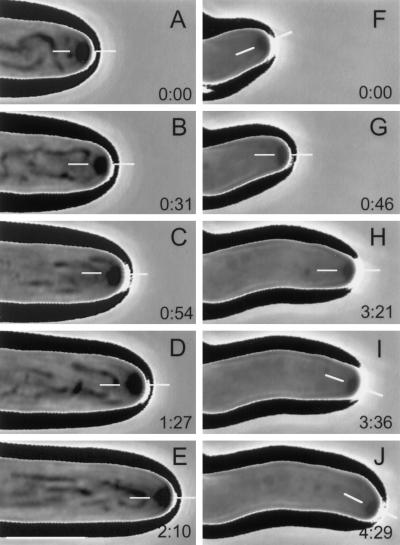
A gene (NhKIN1) encoding a kinesin was cloned from Nectria haematococca genomic DNA by polymerase chain reaction amplification, using primers corresponding to conserved regions of known kinesin-encoding genes. Sequence analysis showed that NhKIN1 belongs to the subfamily of conventional kinesins and is distinct from any of the currently designated kinesin-related protein subfamilies. Deletion of NhKIN1 by transformation-mediated homologous recombination caused several dramatic phenotypes: a 50% reduction in colony growth rate, helical or wavy hyphae with reduced diameter, and subcellular abnormalities including withdrawal of mitochondria from the growing hyphal apex and reduction in the size of the Spitzenkörper, an apical aggregate of secretory vesicles. The effects on mitochondria and Spitzenkörper were not due to altered microtubule distribution, as microtubules were abundant throughout the length of hyphal tip cells of the mutant. The rate of spindle elongation during anaphase B of mitosis was reduced 11%, but the rate was not significantly different from that of wild type. This lack of a substantial mitotic phenotype is consistent with the primary role of the conventional kinesins in organelle motility rather than mitosis. Our results provide further evidence that the microtubule-based motility mechanism has a direct role in apical transport of secretory vesicles and the first evidence for its role in apical transport of mitochondria in a filamentous fungus. They also include a unique demonstration that a microtubule-based motor protein is essential for normal positioning of the Spitzenkörper, thus providing a new insight into the cellular basis for the aberrant hyphal morphology.
Figures

References
-
- Aist JR, Bayles CJ. Video motion analysis of mitotic events in living cells of the fungus Fusarium solani. Cell Motil Cytoskel. 1988;9:325–336.
-
- Aist JR, Bayles CJ. Detection of spindle pushing forces in vivo during anaphase B in the fungus Nectria haematococca. Cell Motil Cytoskel. 1991;19:18–24.
-
- Aist JR, Bayles CJ, Tao W, Berns MW. Direct experimental evidence for the existence, structural basis and function of astral forces during anaphase B in vivo. J Cell Sci. 1991;100:279–288. - PubMed
-
- Allan V. Membrane traffic motors. FEBS Lett. 1995;369:101–106. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
