

Binding of signal recognition particle gives ribosome/nascent chain complexes a competitive advantage in endoplasmic reticulum membrane interaction
- PMID: 9436994
- PMCID: PMC25224
- DOI: 10.1091/mbc.9.1.103
Binding of signal recognition particle gives ribosome/nascent chain complexes a competitive advantage in endoplasmic reticulum membrane interaction
Abstract
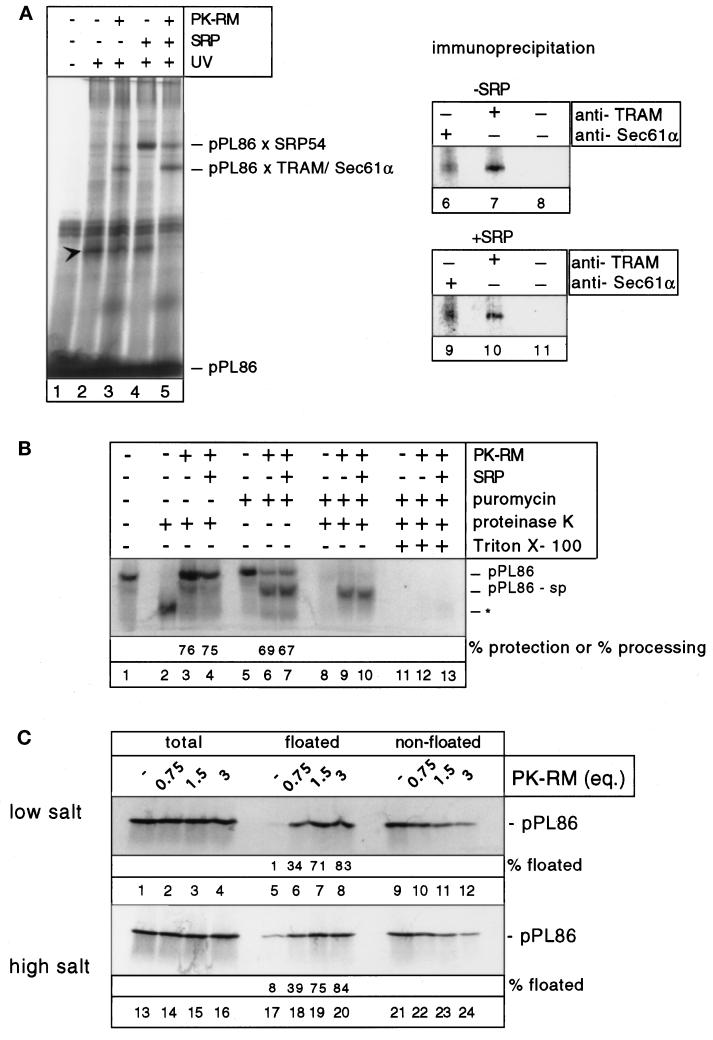
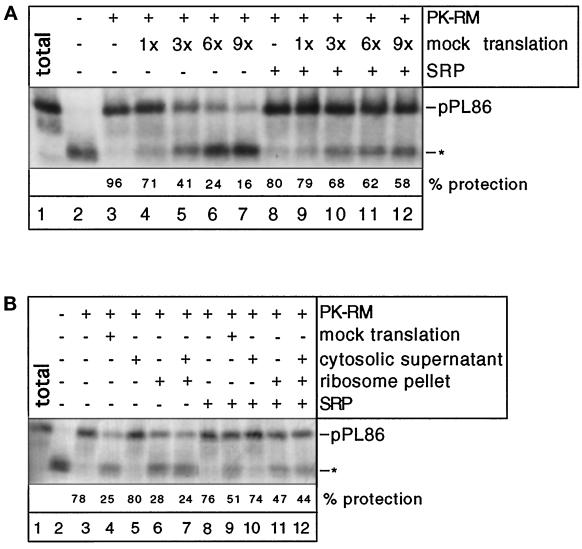
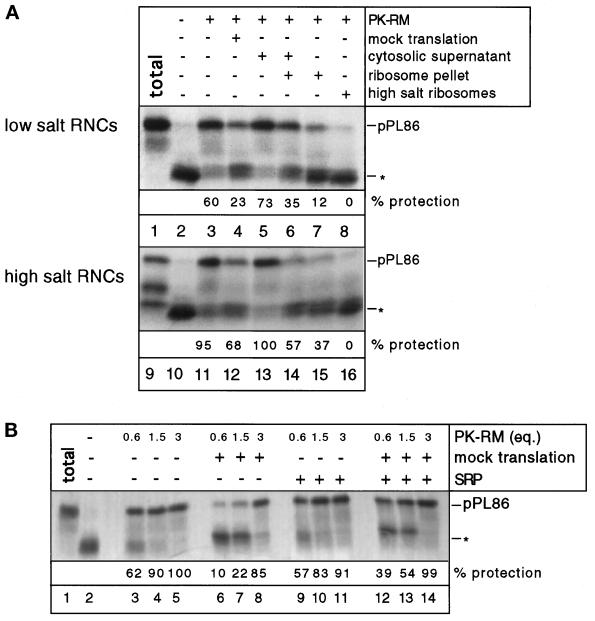
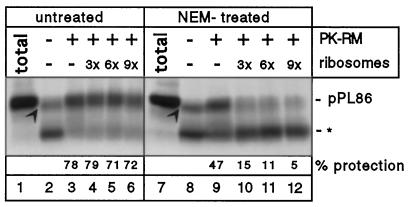
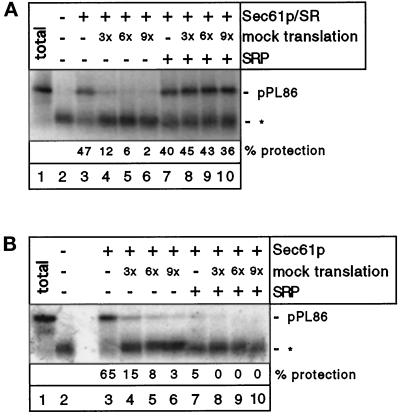
Most secretory and membrane proteins are sorted by signal sequences to the endoplasmic reticulum (ER) membrane early during their synthesis. Targeting of the ribosome-nascent chain complex (RNC) involves the binding of the signal sequence to the signal recognition particle (SRP), followed by an interaction of ribosome-bound SRP with the SRP receptor. However, ribosomes can also independently bind to the ER translocation channel formed by the Sec61p complex. To explain the specificity of membrane targeting, it has therefore been proposed that nascent polypeptide-associated complex functions as a cytosolic inhibitor of signal sequence- and SRP-independent ribosome binding to the ER membrane. We report here that SRP-independent binding of RNCs to the ER membrane can occur in the presence of all cytosolic factors, including nascent polypeptide-associated complex. Nontranslating ribosomes competitively inhibit SRP-independent membrane binding of RNCs but have no effect when SRP is bound to the RNCs. The protective effect of SRP against ribosome competition depends on a functional signal sequence in the nascent chain and is also observed with reconstituted proteoliposomes containing only the Sec61p complex and the SRP receptor. We conclude that cytosolic factors do not prevent the membrane binding of ribosomes. Instead, specific ribosome targeting to the Sec61p complex is provided by the binding of SRP to RNCs, followed by an interaction with the SRP receptor, which gives RNC-SRP complexes a selective advantage in membrane targeting over nontranslating ribosomes.
Figures

Similar articles
-
Signal recognition particle-dependent targeting of ribosomes to the rough endoplasmic reticulum in the absence and presence of the nascent polypeptide-associated complex.Mol Biol Cell. 1998 Jan;9(1):117-30. doi: 10.1091/mbc.9.1.117. Mol Biol Cell. 1998. PMID: 9436995 Free PMC article.
-
The intrinsic ability of ribosomes to bind to endoplasmic reticulum membranes is regulated by signal recognition particle and nascent-polypeptide-associated complex.Proc Natl Acad Sci U S A. 1995 Oct 10;92(21):9435-9. doi: 10.1073/pnas.92.21.9435. Proc Natl Acad Sci U S A. 1995. PMID: 7568149 Free PMC article.
-
Ribosome binding to and dissociation from translocation sites of the endoplasmic reticulum membrane.Mol Biol Cell. 2006 Sep;17(9):3860-9. doi: 10.1091/mbc.e06-05-0439. Epub 2006 Jul 5. Mol Biol Cell. 2006. PMID: 16822833 Free PMC article.
-
Origins and evolution of cotranslational transport to the ER.Adv Exp Med Biol. 2007;607:52-60. doi: 10.1007/978-0-387-74021-8_4. Adv Exp Med Biol. 2007. PMID: 17977458 Review.
-
Structure, function and evolution of the signal recognition particle.EMBO J. 2003 Jul 15;22(14):3479-85. doi: 10.1093/emboj/cdg337. EMBO J. 2003. PMID: 12853463 Free PMC article. Review.
Cited by
-
Characterization of the signal that directs Tom20 to the mitochondrial outer membrane.J Cell Biol. 2000 Oct 16;151(2):277-88. doi: 10.1083/jcb.151.2.277. J Cell Biol. 2000. PMID: 11038175 Free PMC article.
-
The endoplasmic reticulum membrane is permeable to small molecules.Mol Biol Cell. 2004 Feb;15(2):447-55. doi: 10.1091/mbc.e03-05-0325. Epub 2003 Nov 14. Mol Biol Cell. 2004. PMID: 14617815 Free PMC article.
-
A general mechanism for regulation of access to the translocon: competition for a membrane attachment site on ribosomes.Proc Natl Acad Sci U S A. 1998 Nov 10;95(23):13425-30. doi: 10.1073/pnas.95.23.13425. Proc Natl Acad Sci U S A. 1998. PMID: 9811816 Free PMC article.
-
Bioinformatics analysis and alternative polyadenylation in Heat Shock Proteins 70 (HSP70) family members.Int J Physiol Pathophysiol Pharmacol. 2024 Dec 25;16(6):138-151. doi: 10.62347/CWPE7813. eCollection 2024. Int J Physiol Pathophysiol Pharmacol. 2024. PMID: 39850245 Free PMC article.
-
Role of the cytoplasmic segments of Sec61alpha in the ribosome-binding and translocation-promoting activities of the Sec61 complex.J Cell Biol. 2000 Jul 10;150(1):53-64. doi: 10.1083/jcb.150.1.53. J Cell Biol. 2000. PMID: 10893256 Free PMC article.
References
-
- Bacher G, Lütcke H, Jungnickel B, Rapoport TA, Dobberstein B. Regulation by the ribosome of the GTPase of the signal-recognition particle during protein targeting. Nature. 1996;381:248–251. - PubMed
-
- Borgese N, Mok W, Kreibich G, Sabatini DD. Ribosomal-membrane interaction: in vitro binding of ribosomes to microsomal membranes. J Mol Biol. 1974;88:559–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
