

Mitochondrial membrane potential and nuclear changes in apoptosis caused by serum and nerve growth factor withdrawal: time course and modification by (-)-deprenyl
- PMID: 9437015
- PMCID: PMC6792769
- DOI: 10.1523/JNEUROSCI.18-03-00932.1998
Mitochondrial membrane potential and nuclear changes in apoptosis caused by serum and nerve growth factor withdrawal: time course and modification by (-)-deprenyl
Abstract
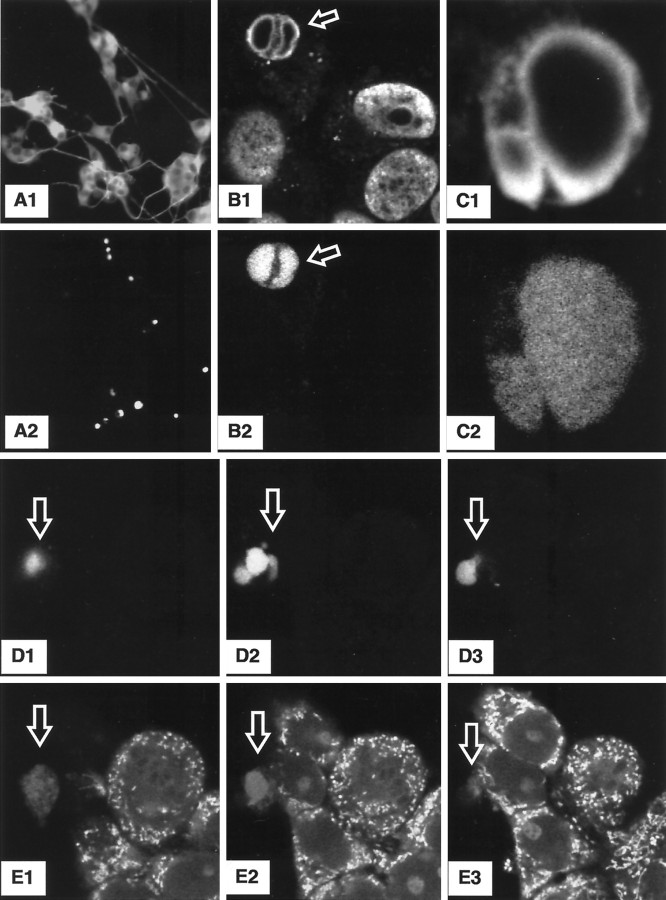
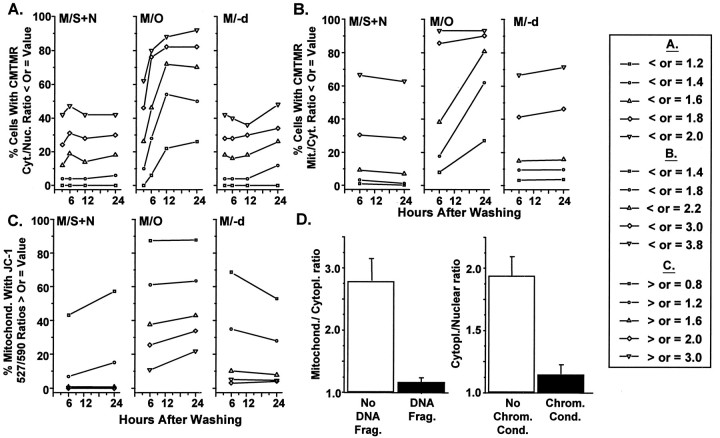
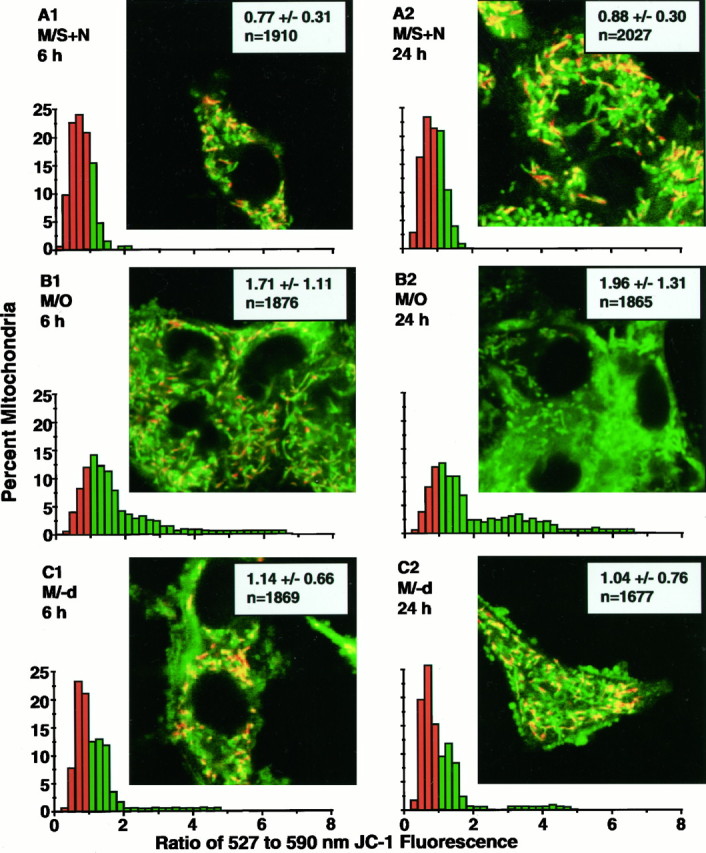
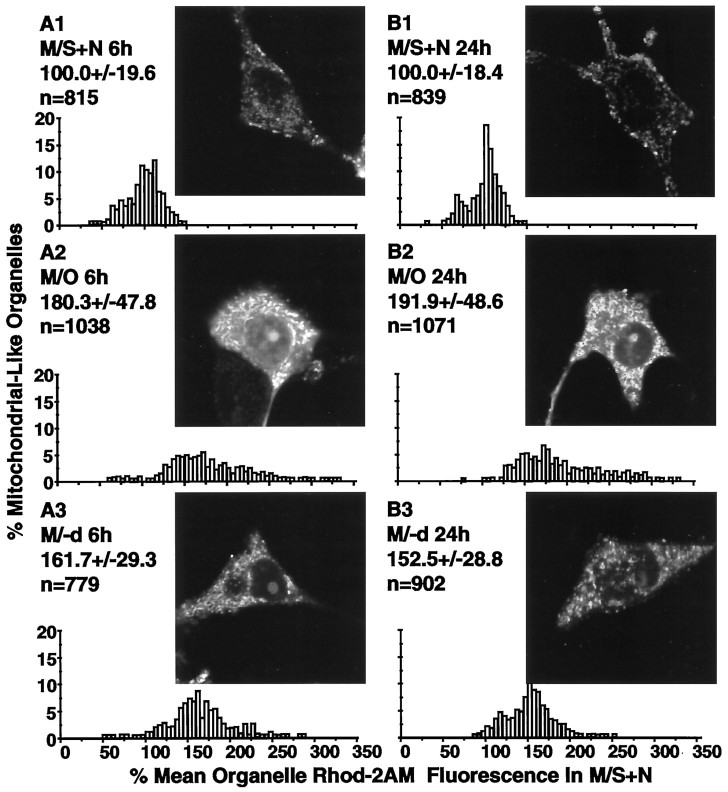
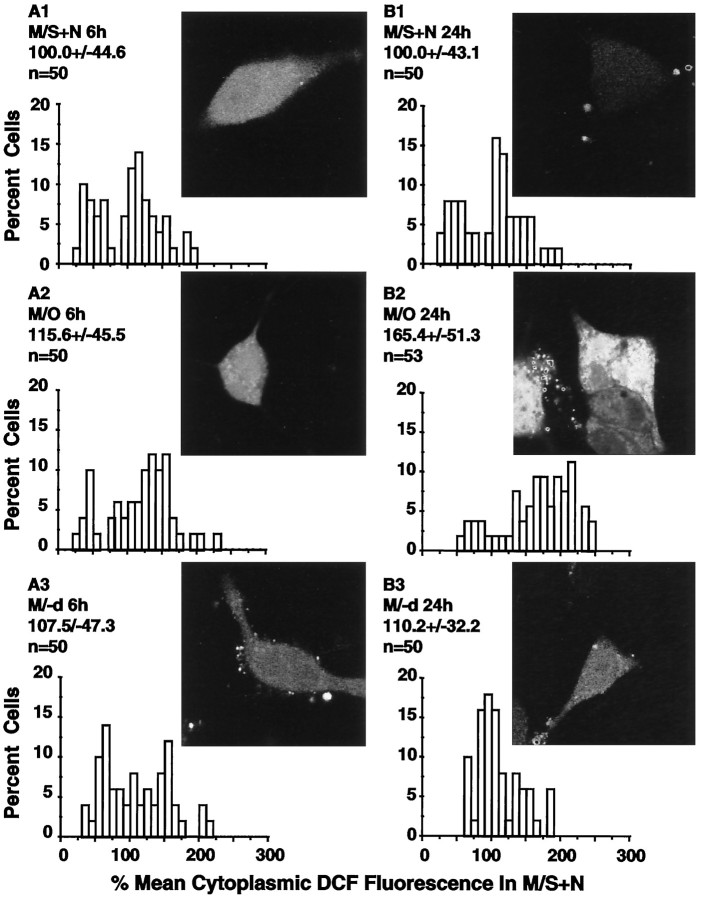
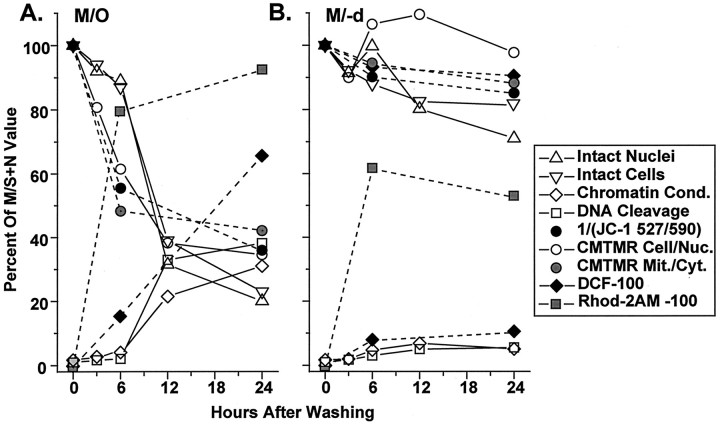
Studies in non-neural cells have suggested that a fall in mitochondrial membrane potential (DeltaPsiM) is one of the earliest events in apoptosis. It is not known whether neural apoptosis caused by nerve growth factor (NGF) and serum withdrawal involves a decrease in DeltaPsiM. We used epifluorescence and laser confocal microscopy with the mitochondrial potentiometric dyes chloromethyl-tetramethylrosamine methyl ester and 5,5',6, 6'-tetrachloro-1,1',3,3'-tetraethybenzimidazol carbocyanine iodide to estimate DeltaPsiM. PC12 cells were differentiated in media containing serum and NGF for 6 d before withdrawal of trophic support. After washing, the cells were incubated with media containing serum and NGF (M/S+N), media without serum and NGF, or media with the "trophic-like" monoamine oxidase B inhibitor, (-)-deprenyl. Mitochondria in cells without trophic support underwent a progressive shift to lower DeltaPsiM values that was significant by 3 hr after washing. The percentages of cells with nuclear chromatin condensation or nuclear DNA fragmentation were not significantly increased above those for cells in M/S+N until 6 hr after washing. Replacement of cells into M/S+N or treatment with (-)-deprenyl markedly reduced the proportion of mitochondria with decreased DeltaPsiM. Measurements of cytoplasmic peroxyl radical levels with 2',7'-dihydrodichlorofluorescein fluorescence and intramitochondrial Ca2+ with dihydro-rhodamine-2-acetylmethyl ester indicated that cytoplasmic peroxyl radical levels were not increased until after 6 hr, whereas increases in intramitochondrial Ca2+ paralleled the decreases in DeltaPsiM. (-)-Deprenyl appeared to alter the relationship between intramitochondrial Ca2+ levels and DeltaPsiM, possibly through its reported capacity to increase the synthesis of proteins such as BCL-2.
Figures
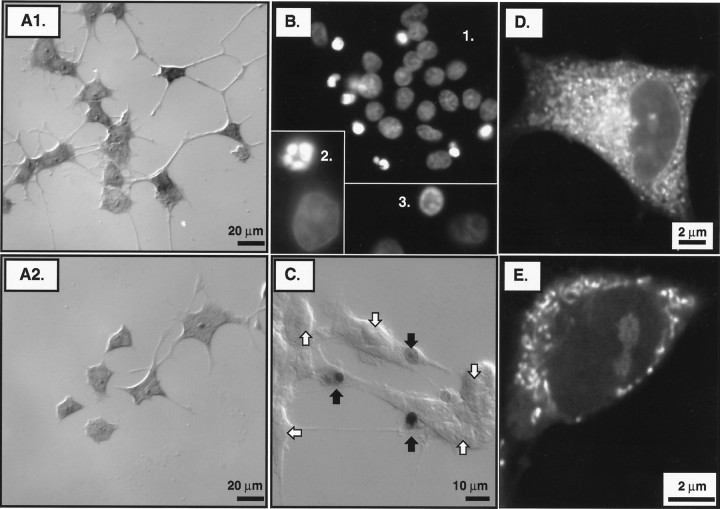
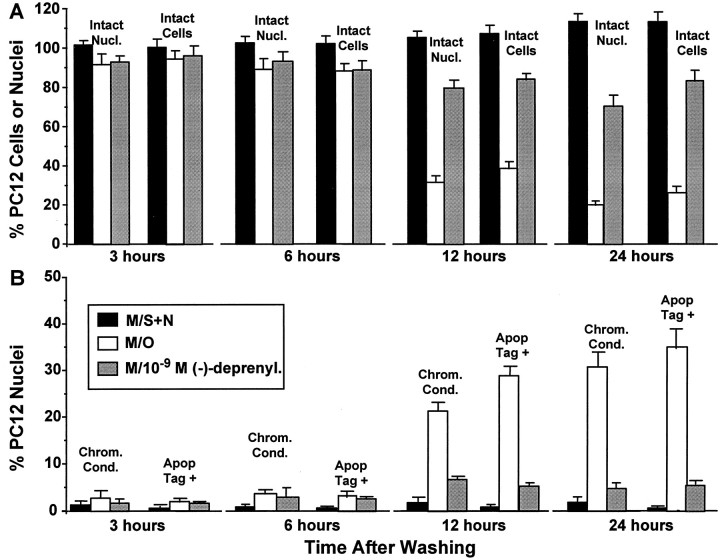
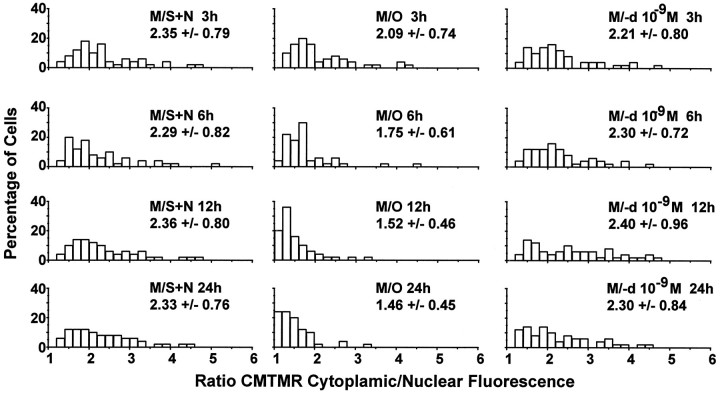
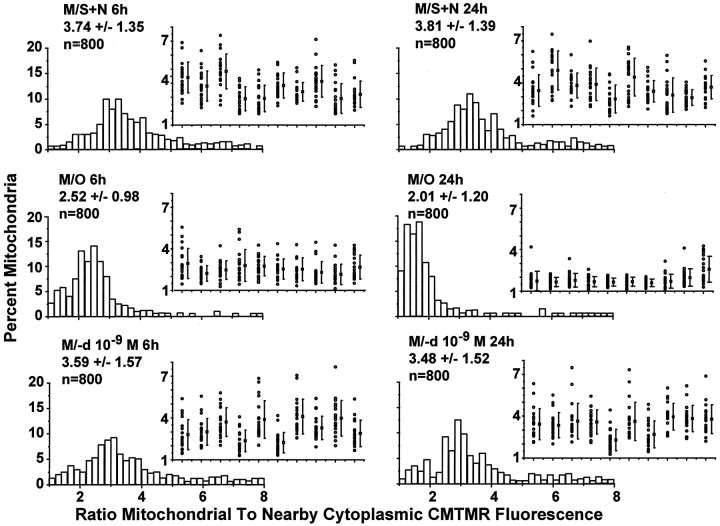
References
-
- Beal MF. Mitochondria, free radicals, and neurodegeneration. Curr Opin Neurobiol. 1996;6:661–666. - PubMed
-
- Biagini G, Zoli M, Fuxe K, Agnati LF. l-Deprenyl increases GFAP immunoreactivity selectively in activated astrocytes in rat brain. NeuroReport. 1993;4:955–958. - PubMed
-
- Biagini G, Frasoldati A, Fuxe K, Agnati LF. The concept of astrocyte-kinetic drug in the treatment of neurodegenerative diseases: evidence for l-deprenyl-induced activation of reactive astrocytes. Neurochem Int. 1994;25:17–22. - PubMed
-
- Calarco PG. Polarization of mitochondria in the fertilized mouse oocyte. Dev Genet. 1995;16:36–43. - PubMed
-
- Carayon P, Portier M, Dussossoy D, Bord A, Petipretre G, Canat X, Le Fur G, Casellas P. Involvement of peripheral benzodiazepine receptors in the protection of hematopoietic cells against oxygen radical damage. Blood. 1996;87:3170–3178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous