

Properties of the P-type ATPases encoded by the copAP operons of Helicobacter pylori and Helicobacter felis
- PMID: 9440521
- PMCID: PMC106887
- DOI: 10.1128/JB.180.2.317-329.1998
Properties of the P-type ATPases encoded by the copAP operons of Helicobacter pylori and Helicobacter felis
Abstract
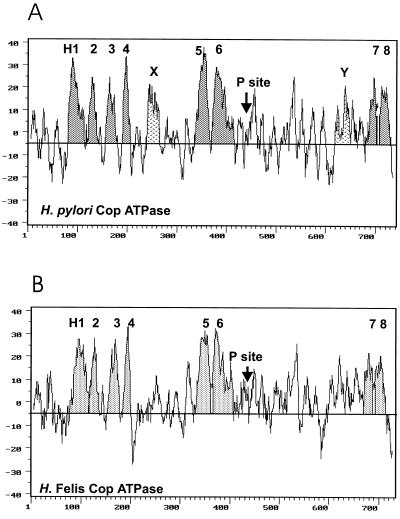
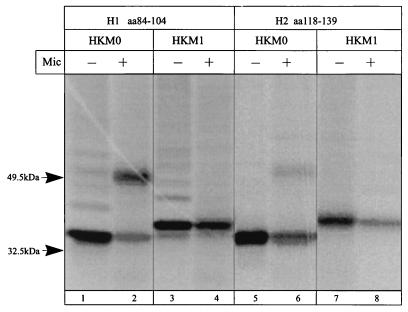
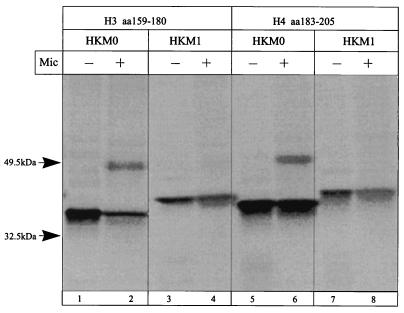
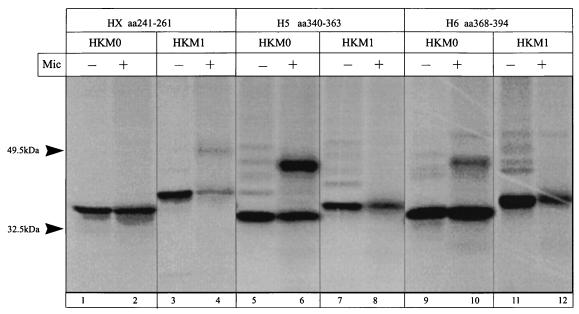
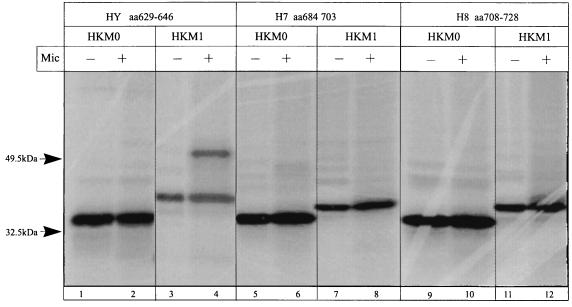
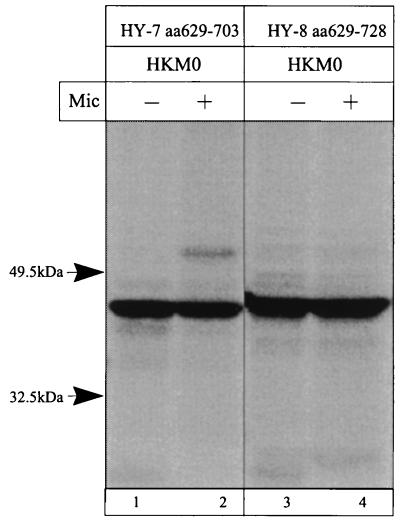
The cop operons of Helicobacter pylori and Helicobacter felis were cloned by gene library screening. Both operons contain open reading frames for a P-type ion pump (CopA) with homology to Cd2+ and Cu2+ ATPases and a putative ion binding protein (CopP), the latter representing a CopZ homolog of the copYZAB operon of Enterococcus hirae. The predicted CopA ATPases contained an N-terminal GMXCXXC ion binding motif and a membrane-associated CPC sequence. A synthetic N-terminal peptide of the H. pylori CopA ATPase bound to Cu2+ specifically, and gene disruption mutagenesis of CopA resulted in an enhanced growth sensitivity of H. pylori to Cu2+ but not to other divalent cations. As determined experimentally, H. pylori CopA contains four pairs of transmembrane segments (H1 to H8), with the ATP binding and phosphorylation domains lying between H6 and H7, as found for another putative transition metal pump of H. pylori (K. Melchers, T. Weitzenegger, A. Buhmann, W. Steinhilber, G. Sachs, and K. P. Schäfer, J. Biol. Chem. 271:446-457, 1996). The corresponding transmembrane segments of the H. felis CopA pump were identified by hydrophobicity analysis and via sequence similarity. To define functional domains, similarly oriented regions of the two enzymes were examined for sequence identity. Regions with high degrees of identity included the N-terminal Cu2+ binding domain, the regions of ATP binding and phosphorylation in the energy transduction domain, and a transport domain consisting of the last six transmembrane segments with conserved cysteines in H4, H6, and H7. The data suggest that H. pylori and H. felis employ conserved mechanisms of ATPase-dependent copper resistance.
Figures


Similar articles
-
Cloning and membrane topology of a P type ATPase from Helicobacter pylori.J Biol Chem. 1996 Jan 5;271(1):446-57. doi: 10.1074/jbc.271.1.446. J Biol Chem. 1996. PMID: 8550601
-
Membrane topology of CadA homologous P-type ATPase of Helicobacter pylori as determined by expression of phoA fusions in Escherichia coli and the positive inside rule.Res Microbiol. 1999 Oct;150(8):507-20. doi: 10.1016/s0923-2508(99)00106-0. Res Microbiol. 1999. PMID: 10577484
-
Properties and function of the P type ion pumps cloned from Helicobacter pylori.Acta Physiol Scand Suppl. 1998 Aug;643:123-35. Acta Physiol Scand Suppl. 1998. PMID: 9789554
-
The structure and function of heavy metal transport P1B-ATPases.Biometals. 2007 Jun;20(3-4):233-48. doi: 10.1007/s10534-006-9055-6. Epub 2007 Jan 12. Biometals. 2007. PMID: 17219055 Review.
-
Copper homeostasis in Enterococcus hirae.FEMS Microbiol Rev. 2003 Jun;27(2-3):183-95. doi: 10.1016/S0168-6445(03)00053-6. FEMS Microbiol Rev. 2003. PMID: 12829267 Review.
Cited by
-
Emergence of diverse Helicobacter species in the pathogenesis of gastric and enterohepatic diseases.Clin Microbiol Rev. 2001 Jan;14(1):59-97. doi: 10.1128/CMR.14.1.59-97.2001. Clin Microbiol Rev. 2001. PMID: 11148003 Free PMC article. Review.
-
The construction of a new integrative vector with a new selective marker of copper resistance for glycerol producer Candida glycerinogenes.Curr Microbiol. 2012 Apr;64(4):357-64. doi: 10.1007/s00284-011-0075-2. Epub 2012 Jan 12. Curr Microbiol. 2012. PMID: 22237983
-
The high copper tolerance of Candida albicans is mediated by a P-type ATPase.Proc Natl Acad Sci U S A. 2000 Mar 28;97(7):3520-5. doi: 10.1073/pnas.97.7.3520. Proc Natl Acad Sci U S A. 2000. PMID: 10737803 Free PMC article.
-
A Multicopper oxidase (Cj1516) and a CopA homologue (Cj1161) are major components of the copper homeostasis system of Campylobacter jejuni.J Bacteriol. 2008 Dec;190(24):8075-85. doi: 10.1128/JB.00821-08. Epub 2008 Oct 17. J Bacteriol. 2008. PMID: 18931123 Free PMC article.
-
Functional and expression analyses of the cop operon, required for copper resistance in Agrobacterium tumefaciens.J Bacteriol. 2009 Aug;191(16):5159-68. doi: 10.1128/JB.00384-09. Epub 2009 Jun 5. J Bacteriol. 2009. PMID: 19502402 Free PMC article.
References
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1994.
-
- Bamberg K, Sachs G. Topological analysis of the H,K-ATPase using in vitro translation. J Biol Chem. 1994;269:16909–16919. - PubMed
-
- Bayer E, Rapp W. New polymer supports for solid-liquid-phase peptide synthesis. Chem Pept Proteins. 1986;3:3–8.
-
- Bayle D, Weeks D, Sachs G. The membrane topology of the rat sarcoplasmic and endoplasmic reticulum calcium ATPases by in vitro translation scanning. J Biol Chem. 1995;270:25678–25684. - PubMed
-
- Bull P C, Thomas G R, Rommens J M, Forbes J R, Cox D W. The Wilson disease gene is a putative copper transporting P-type ATPase similar to the Menkes gene. Nat Genet. 1993;5:327–336. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
