

Identification, genomic organization, and analysis of the group III capsular polysaccharide genes kpsD, kpsM, kpsT, and kpsE from an extraintestinal isolate of Escherichia coli (CP9, O4/K54/H5)
- PMID: 9440523
- PMCID: PMC106889
- DOI: 10.1128/JB.180.2.338-349.1998
Identification, genomic organization, and analysis of the group III capsular polysaccharide genes kpsD, kpsM, kpsT, and kpsE from an extraintestinal isolate of Escherichia coli (CP9, O4/K54/H5)
Abstract
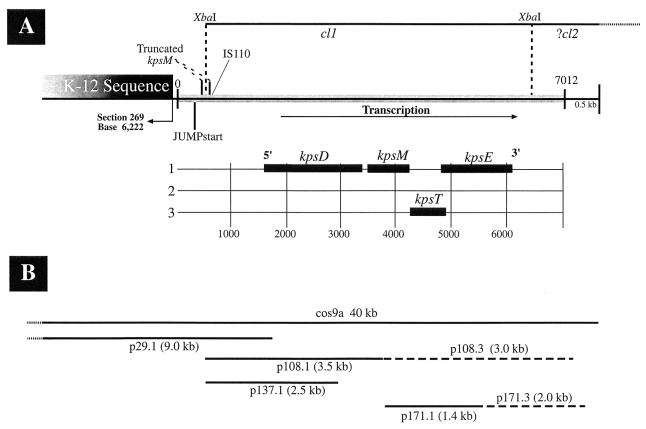
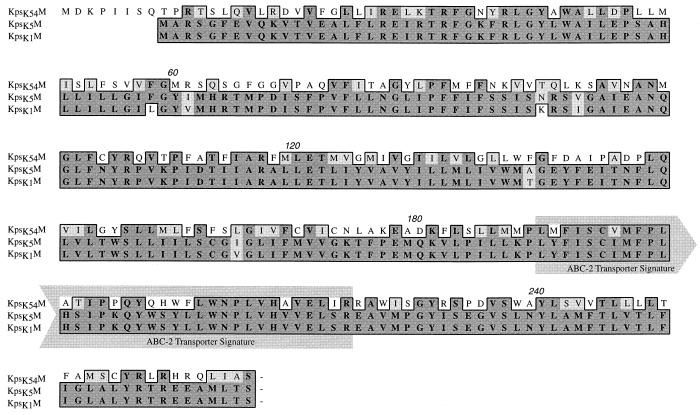
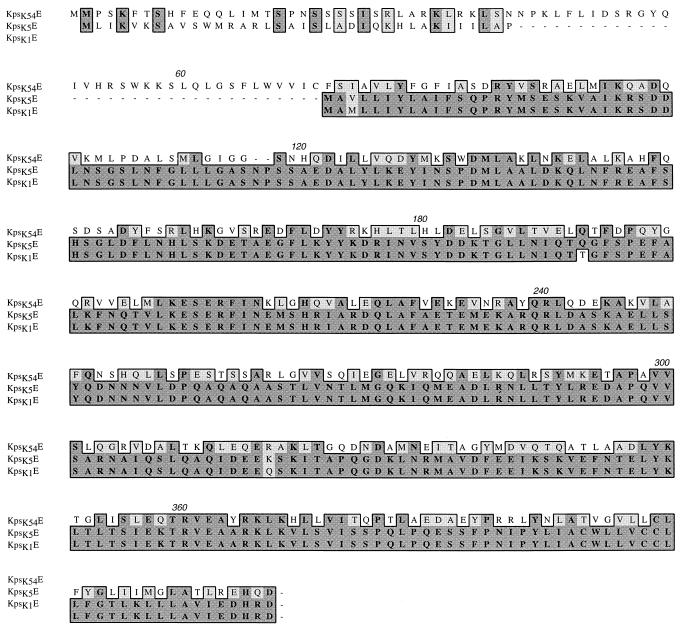
Group III capsular polysaccharides (e.g., K54) of extraintestinal isolates of Escherichia coli, similar to group II capsules (e.g., K1), are important virulence traits that confer resistance to selected host defense components in vitro and potentiate systemic infection in vivo. The genomic organization of group II capsule gene clusters has been established as a serotype-specific region 2 flanked by regions 1 and 3, which contain transport genes that are highly homologous between serotypes. In contrast, the organization of group III capsule gene clusters is not well understood. However, they are defined in part by an absence of genes with significant nucleotide homology to group II capsule transport genes in regions 1 and 3. Evaluation of isogenic, TnphoA-generated, group III capsule-minus derivatives of a clinical blood isolate (CP9, O4/K54/H5) has led to the identification of homologs of the group II capsule transport genes kpsDMTE. These genes and their surrounding regions were sequenced and analyzed. The genomic organization of these genes is distinctly different from that of their group II counterparts. Although kps(K54)DMTE are significantly divergent from their group II homologs at both the DNA and protein levels phoA fusions and computer-assisted analyses suggest that their structures and functions are similar. The putative proteins Kps(K54)M and Kps(K54)T appear to be the integral membrane component and the peripheral ATP-binding component of the ABC-2 transporter family, respectively. The putative Kps(K54)E possesses features similar to those of the membrane fusion protein family that facilitates the passage of large molecules across the periplasm. At one boundary of the capsule gene cluster, a truncated kpsM (kpsM(truncated) and its 5' noncoding regulatory sequence were identified. In contrast to the complete kps(K54)M, this region was highly homologous to the group II kpsM. Fifty-three base pairs 3' from the end of kpsM(truncated) was a sequence 75% homologous to the 39-bp inverted repeat in the IS110 insertion element from Streptomyces coelicolor. Southern analysis established that two copies of this element are present in CP9. These findings are consistent with the hypothesis that CP9 previously possessed group II capsule genes and acquired group III capsule genes via IS110-mediated horizontal transfer.
Figures





Similar articles
-
An extraintestinal, pathogenic isolate of Escherichia coli (O4/K54/H5) can produce a group 1 capsule which is divergently regulated from its constitutively produced group 2, K54 capsular polysaccharide.J Bacteriol. 1993 Dec;175(23):7617-23. doi: 10.1128/jb.175.23.7617-7623.1993. J Bacteriol. 1993. PMID: 8244930 Free PMC article.
-
Cloning and analysis of gene clusters for production of the Escherichia coli K10 and K54 antigens: identification of a new group of serA-linked capsule gene clusters.J Bacteriol. 1995 Jul;177(14):3992-7. doi: 10.1128/jb.177.14.3992-3997.1995. J Bacteriol. 1995. PMID: 7608072 Free PMC article.
-
The construction and characterization of colanic acid deficient mutants in an extraintestinal isolate of Escherichia coli (O4/K54/H5).Microb Pathog. 1995 Apr;18(4):269-78. doi: 10.1016/s0882-4010(05)80003-8. Microb Pathog. 1995. PMID: 7476092
-
Identification of two genes, kpsM and kpsT, in region 3 of the polysialic acid gene cluster of Escherichia coli K1.J Bacteriol. 1991 Aug;173(15):4603-10. doi: 10.1128/jb.173.15.4603-4610.1991. J Bacteriol. 1991. PMID: 1856162 Free PMC article.
-
Coating the surface: a model for expression of capsular polysialic acid in Escherichia coli K1.Mol Microbiol. 1996 Jul;21(2):221-31. doi: 10.1046/j.1365-2958.1996.6461357.x. Mol Microbiol. 1996. PMID: 8858578 Review.
Cited by
-
Canine feces as a reservoir of extraintestinal pathogenic Escherichia coli.Infect Immun. 2001 Mar;69(3):1306-14. doi: 10.1128/IAI.69.3.1306-1314.2001. Infect Immun. 2001. PMID: 11179292 Free PMC article.
-
An ST131 clade and a phylogroup A clade bearing an O101-like O-antigen cluster predominate among bloodstream Escherichia coli isolates from South-West Nigeria hospitals.Microb Genom. 2022 Dec;8(12):mgen000863. doi: 10.1099/mgen.0.000863. Microb Genom. 2022. PMID: 36748556 Free PMC article.
-
A killed, genetically engineered derivative of a wild-type extraintestinal pathogenic E. coli strain is a vaccine candidate.Vaccine. 2007 May 10;25(19):3859-70. doi: 10.1016/j.vaccine.2007.01.100. Epub 2007 Feb 5. Vaccine. 2007. PMID: 17306426 Free PMC article.
-
Detection and identification of Actinobacillus pleuropneumoniae serotypes 1, 2, and 8 by multiplex PCR.J Clin Microbiol. 2004 Sep;42(9):4344-8. doi: 10.1128/JCM.42.9.4344-4348.2004. J Clin Microbiol. 2004. PMID: 15365041 Free PMC article.
-
The complete genome sequence of Cupriavidus metallidurans strain CH34, a master survivalist in harsh and anthropogenic environments.PLoS One. 2010 May 5;5(5):e10433. doi: 10.1371/journal.pone.0010433. PLoS One. 2010. PMID: 20463976 Free PMC article.
References
-
- Ames G F-L, Mimura C S, Holbrook S R, Shyamala V. Traffic ATPases: a superfamily of transport proteins operating from Escherichia coli to humans. Adv Enzymol. 1992;65:1–47. - PubMed
-
- Bliss J M, Silver R P. Coating the surface: a model for expression of capsular polysialic acid in Escherichia coli K1. Mol Microbiol. 1996;21:221–231. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
