

Identification of a preinitiation step in DNA replication that is independent of origin recognition complex and cdc6, but dependent on cdk2
- PMID: 9442103
- PMCID: PMC2132576
- DOI: 10.1083/jcb.140.2.271
Identification of a preinitiation step in DNA replication that is independent of origin recognition complex and cdc6, but dependent on cdk2
Abstract
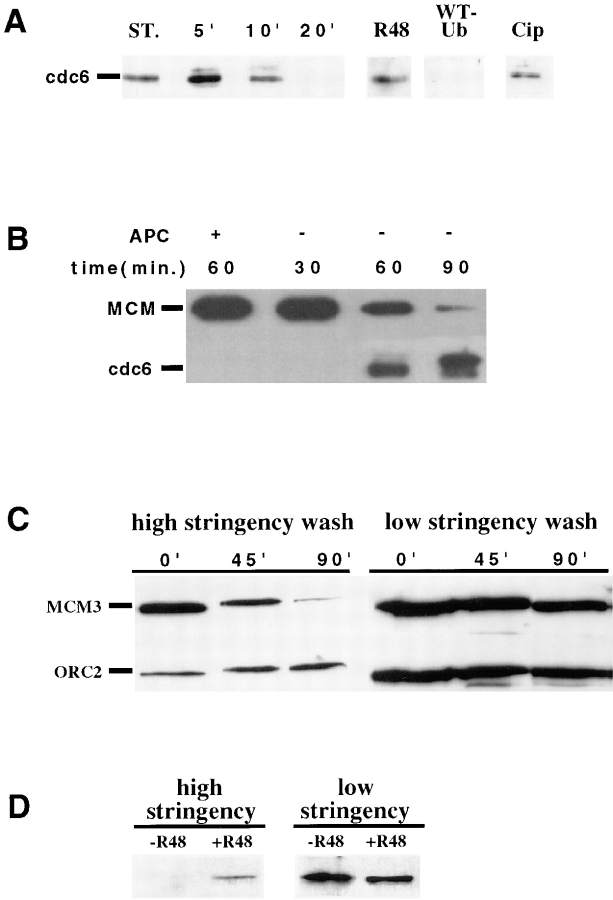
Before initiation of DNA replication, origin recognition complex (ORC) proteins, cdc6, and minichromosome maintenance (MCM) proteins bind to chromatin sequentially and form preinitiation complexes. Using Xenopus laevis egg extracts, we find that after the formation of these complexes and before initiation of DNA replication, cdc6 is rapidly removed from chromatin, possibly degraded by a cdk2-activated, ubiquitin-dependent proteolytic pathway. If this displacement is inhibited, DNA replication fails to initiate. We also find that after assembly of MCM proteins into preinitiation complexes, removal of the ORC from DNA does not block the subsequent initiation of replication. Importantly, under conditions in which both ORC and cdc6 protein are absent from preinitiation complexes, DNA replication is still dependent on cdk2 activity. Therefore, the final steps in the process leading to initiation of DNA replication during S phase of the cell cycle are independent of ORC and cdc6 proteins, but dependent on cdk2 activity.
Figures







Similar articles
-
Cyclin A-dependent kinase activity affects chromatin binding of ORC, Cdc6, and MCM in egg extracts of Xenopus laevis.Eur J Biochem. 1999 Sep;264(2):415-26. doi: 10.1046/j.1432-1327.1999.00613.x. Eur J Biochem. 1999. PMID: 10491086
-
Cyclin E uses Cdc6 as a chromatin-associated receptor required for DNA replication.J Cell Biol. 2001 Mar 19;152(6):1267-78. doi: 10.1083/jcb.152.6.1267. J Cell Biol. 2001. PMID: 11257126 Free PMC article.
-
MCM2-7 complexes bind chromatin in a distributed pattern surrounding the origin recognition complex in Xenopus egg extracts.J Biol Chem. 2002 Sep 6;277(36):33049-57. doi: 10.1074/jbc.M204438200. Epub 2002 Jun 26. J Biol Chem. 2002. PMID: 12087101
-
Cyclin-dependent kinases and S phase control in mammalian cells.Cell Cycle. 2003 Jul-Aug;2(4):316-24. Cell Cycle. 2003. PMID: 12851482 Review.
-
Control of DNA replication licensing in a cell cycle.Genes Cells. 2002 Jun;7(6):523-34. doi: 10.1046/j.1365-2443.2002.00544.x. Genes Cells. 2002. PMID: 12059957 Review.
Cited by
-
Genome wide comparative comprehensive analysis of Plasmodium falciparum MCM family with human host.Commun Integr Biol. 2012 Nov 1;5(6):607-15. doi: 10.4161/cib.21759. Commun Integr Biol. 2012. PMID: 23336032 Free PMC article.
-
PD-1 inhibits T cell proliferation by upregulating p27 and p15 and suppressing Cdc25A.Cell Cycle. 2012 Dec 1;11(23):4305-9. doi: 10.4161/cc.22135. Epub 2012 Oct 3. Cell Cycle. 2012. PMID: 23032366 Free PMC article.
-
A requirement for MCM7 and Cdc45 in chromosome unwinding during eukaryotic DNA replication.EMBO J. 2004 Sep 15;23(18):3667-76. doi: 10.1038/sj.emboj.7600369. Epub 2004 Aug 26. EMBO J. 2004. PMID: 15329670 Free PMC article.
-
Xenopus Cdc6 performs separate functions in initiating DNA replication.Mol Biol Cell. 2002 Apr;13(4):1298-312. doi: 10.1091/mbc.01-08-0382. Mol Biol Cell. 2002. PMID: 11950940 Free PMC article.
-
Cdc45 limits replicon usage from a low density of preRCs in mammalian cells.PLoS One. 2011 Mar 1;6(3):e17533. doi: 10.1371/journal.pone.0017533. PLoS One. 2011. PMID: 21390258 Free PMC article.
References
-
- Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature. 1992;357:128–134. - PubMed
-
- Broek D, Bartlett R, Crawford K, Nurse P. Involvement of p34cdc2 in establishing the dependency of S phase on mitosis. Nature. 1991;349:388–393. - PubMed
-
- Carpenter PB, Mueller PR, Dunphy WG. Role for a XenopusOrc2-related protein in controlling DNA replication. Nature. 1996;379:357–360. - PubMed
-
- Chau V, Tobias JW, Bachmair A, Marriott D, Ecker DJ, Gonda DK, Varshavsky A. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science. 1989;243:1576–1583. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
