

Bone morphogenetic protein signaling is required for maintenance of differentiated phenotype, control of proliferation, and hypertrophy in chondrocytes
- PMID: 9442116
- PMCID: PMC2132568
- DOI: 10.1083/jcb.140.2.409
Bone morphogenetic protein signaling is required for maintenance of differentiated phenotype, control of proliferation, and hypertrophy in chondrocytes
Abstract
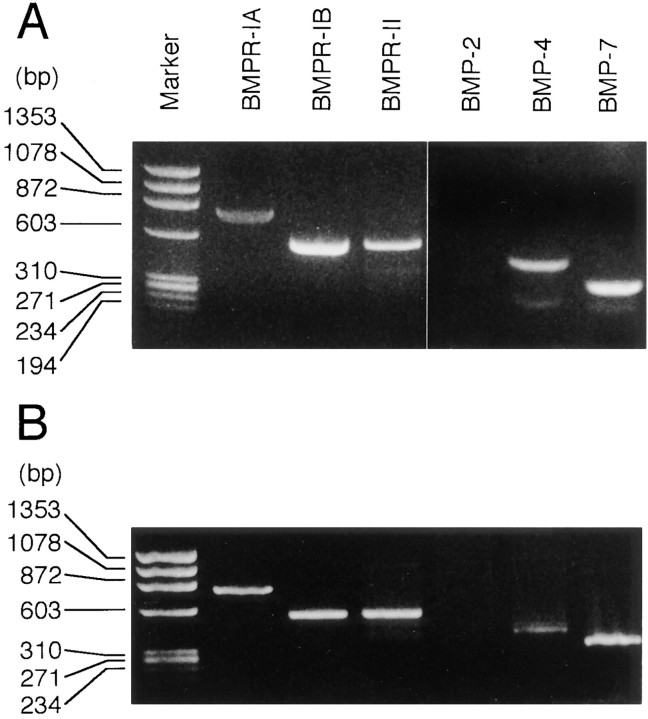
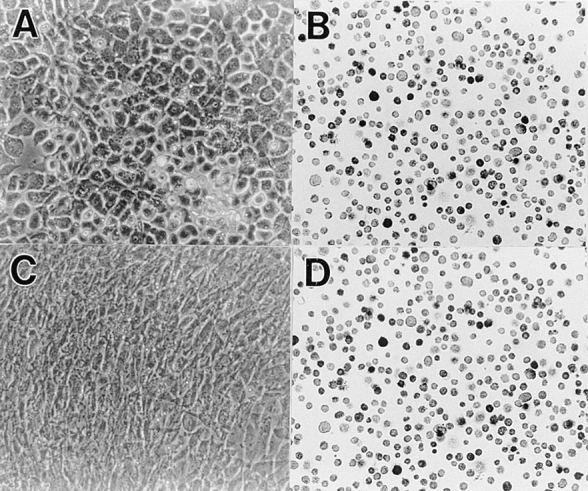
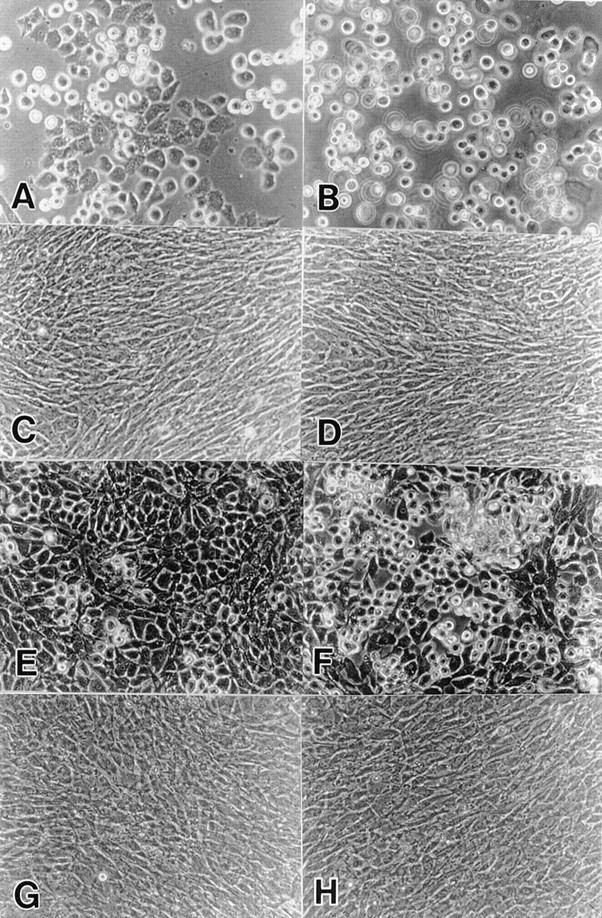
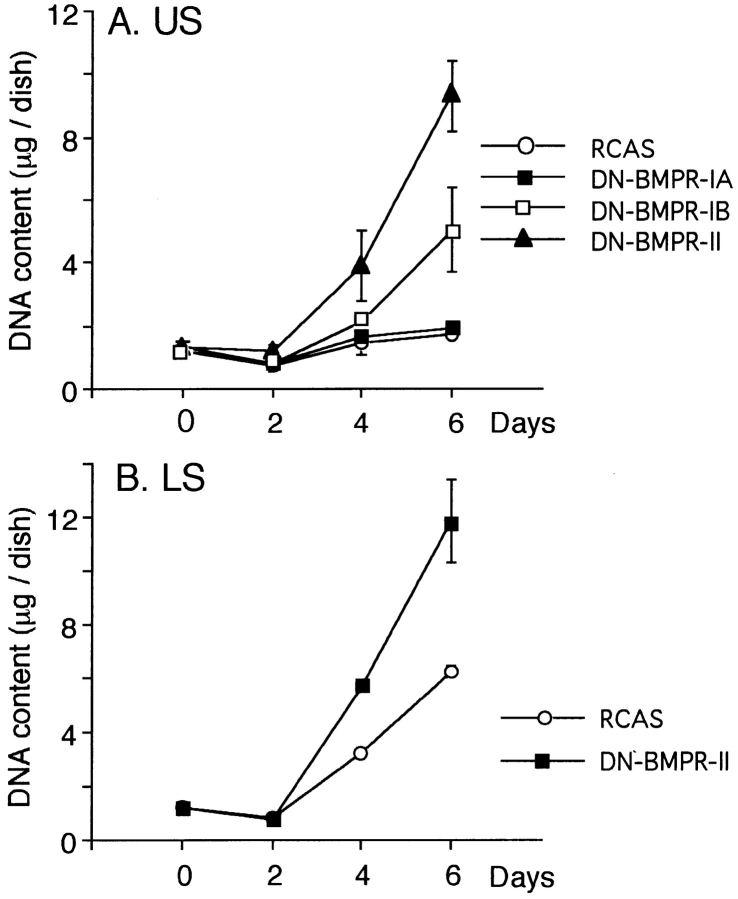
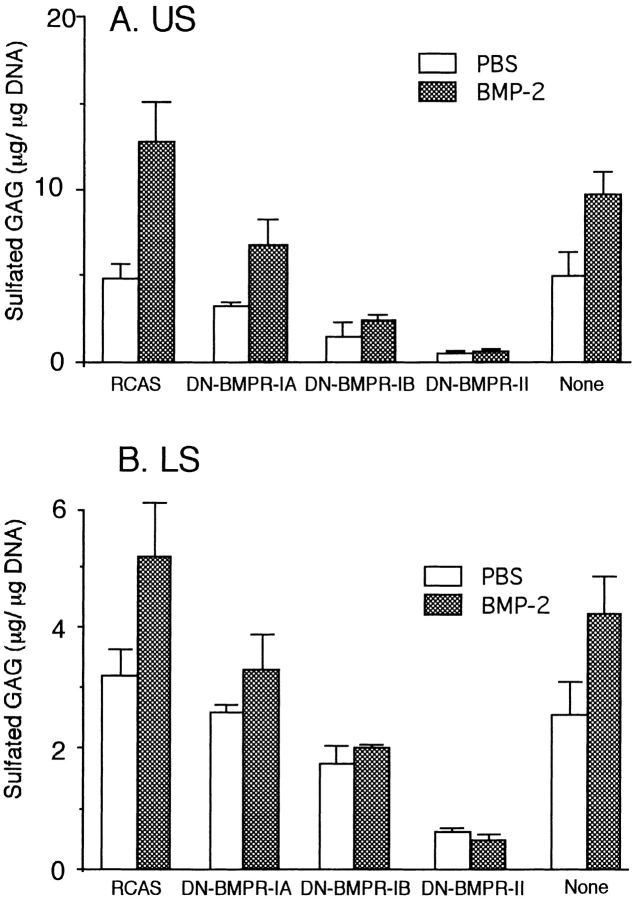
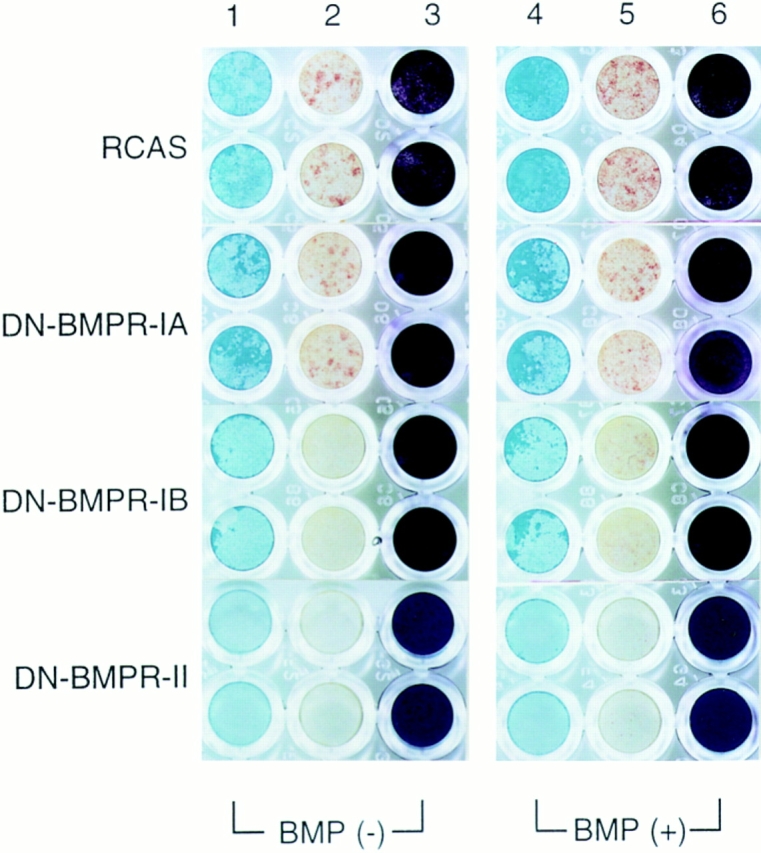
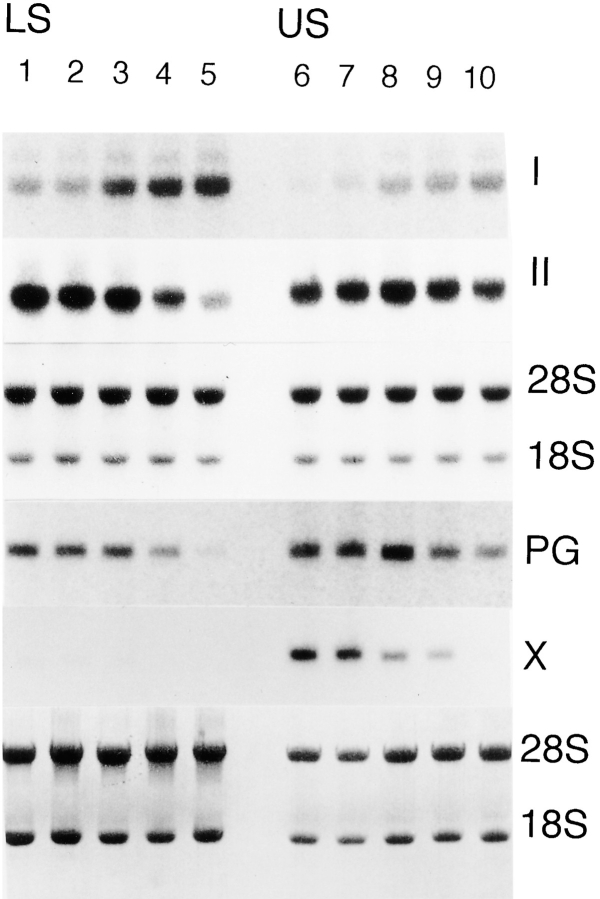
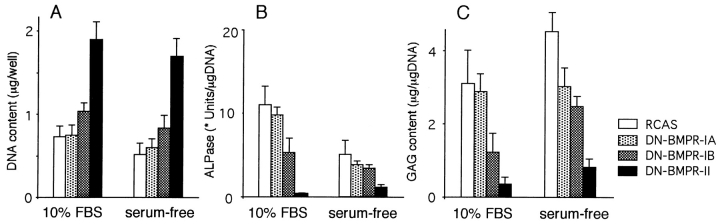
To examine the role of bone morphogenetic protein (BMP) signaling in chondrocytes during endochondral ossification, the dominant negative (DN) forms of BMP receptors were introduced into immature and mature chondrocytes isolated from lower and upper portions of chick embryo sternum, respectively. We found that control sternal chondrocyte populations expressed type IA, IB, and II BMP receptors as well as BMP-4 and -7. Expression of a DN-type II BMP receptor (termed DN-BMPR-II) in immature lower sternal (LS) chondrocytes led to a loss of differentiated functions; compared with control cells, the DN-BMPR- II-expressing LS chondrocytes proliferated more rapidly, acquired a fibroblastic morphology, showed little expression of type II collagen and aggrecan genes, and upregulated type I collagen gene expression. Expression of DN-BMPR-II in mature hypertrophic upper sternal (US) chondrocytes caused similar effects. In addition, the DN-BMPR-II-expressing US cells exhibited little alkaline phosphatase activity and type X collagen gene expression, while the control US cells produced both alkaline phosphatase and type X collagen. Both DN-BMPR-II-expressing US and LS chondrocytes failed to respond to treatment with BMP-2 . When we examined the effects of DN forms of types IA and IB BMP receptors, we found that DN-BMPR-IA had little effect, while DN-BMPR-IB had similar but weaker effects compared with those of DN-BMPR-II. We conclude that BMP signaling, particularly that mediated by the type II BMP receptor, is required for maintenance of the differentiated phenotype, control of cell proliferation, and expression of hypertrophic phenotype.
Figures

References
-
- Benya PD, Shaffer JD. Dedifferentiated chondrocytes reexpress the differentiated collagen phenotype when cultured in agarose gels. Cell. 1982;30:215–224. - PubMed
-
- Bessey OA, Lowry OH, Brock MJ. A method for the rapid determination of alkaline phosphatase with five cubic millimeters of serum. J Biol Chem. 1946;164:321–329. - PubMed
-
- Carey DE, Liu X. Expression of bone morphogenetic protein-6 messenger RNA in bovine growth plate chondrocytes of different size. J Bone Miner Res. 1995;10:401–405. - PubMed
-
- Chen P, Carrington JL, Hammonds RG, Reddi AH. Stimulation of chondrogenesis in limb bud mesoderm cells by recombinant human bone morphogenetic protein 2B (BMP-2B) and modulation by transforming growth factor β1 and β2. Exp Cell Res. 1991;195:509–515. - PubMed
-
- Dewulf N, Verschueren K, Lonnoy O, Moren A, Grimsby S, Vande-Spiegle K, Miyazono K, Huylebroeck D, Ten-Dijke P. Distinct spatial and temporal expression patterns of two type I receptors for bone morphogenetic proteins during mouse embryogenesis. Endocrinology. 1995;136:2652–2653. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
