

Measles viruses with altered envelope protein cytoplasmic tails gain cell fusion competence
- PMID: 9445022
- PMCID: PMC124600
- DOI: 10.1128/JVI.72.2.1224-1234.1998
Measles viruses with altered envelope protein cytoplasmic tails gain cell fusion competence
Abstract
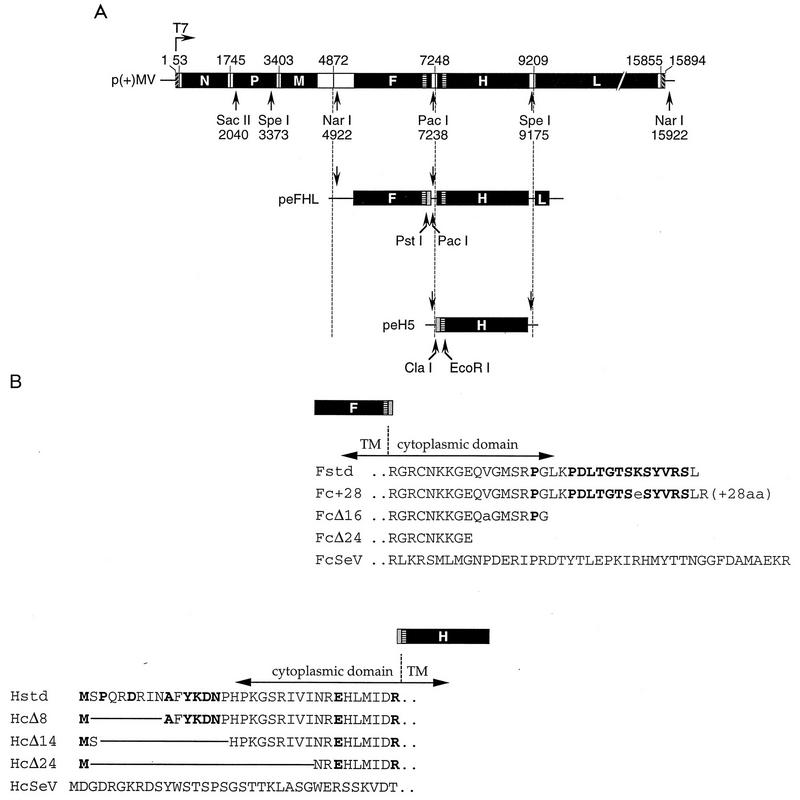
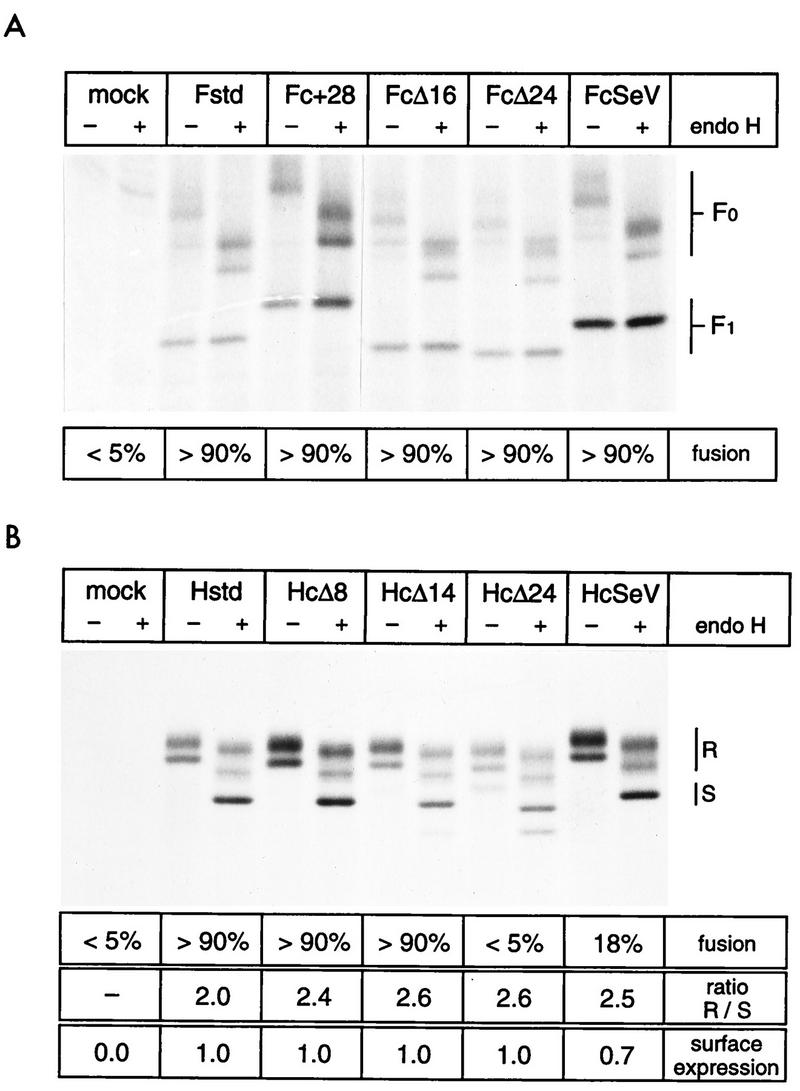
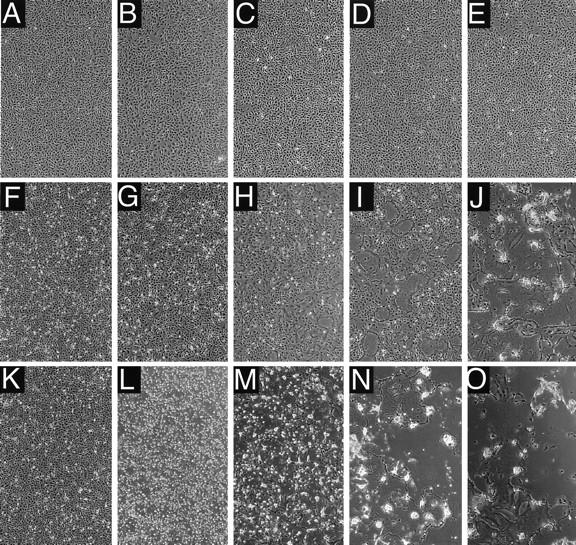
The cytoplasmic tail of the measles virus (MV) fusion (F) protein is often altered in viruses which spread through the brain of patients suffering from subacute sclerosing panencephalitis (SSPE). We transferred the coding regions of F tails from SSPE viruses in an MV genomic cDNA. Similarly, we constructed and transferred mutated tail-encoding regions of the other viral glycoprotein hemagglutinin (H) gene. From the mutated genomic cDNAs, we achieved rescue of viruses that harbor different alterations of the F tail, deletions in the membrane-distal half of the H tail, and combinations of these mutations. Viruses with alterations in any of the tails spread rapidly through the monolayer via enhanced cell-cell fusion. Double-tail mutants had even higher fusion competence but slightly decreased infectivity. Analysis of the protein composition of released mutant viral particles indicated that the tails are necessary for accurate virus envelope assembly and suggested a direct F tail-matrix (M) protein interaction. Since even tail-altered glycoproteins colocalized with M protein in intracellular patches, additional interactions may exist. We conclude that in MV infections, including SSPE, the glycoprotein tails are involved not only in virus envelope assembly but also in the control of virus-induced cell fusion.
Figures
References
-
- Billeter M A, Cattaneo R. Molecular biology of defective measles virus persisting in the human central nervous system. In: Kingsbury D W, editor. The paramyxoviruses. New York, N.Y: Plenum Press; 1991. pp. 323–345.
-
- Buechi M, Bächi T. Microscopy of internal structures of Sendai virus associated with the cytoplasmic surface of host membranes. Virology. 1982;120:349–359. - PubMed
-
- Cathomen, T. 1991. Unpublished data.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
