

The representation of the ipsilateral visual field in human cerebral cortex
- PMID: 9448246
- PMCID: PMC33803
- DOI: 10.1073/pnas.95.3.818
The representation of the ipsilateral visual field in human cerebral cortex
Abstract
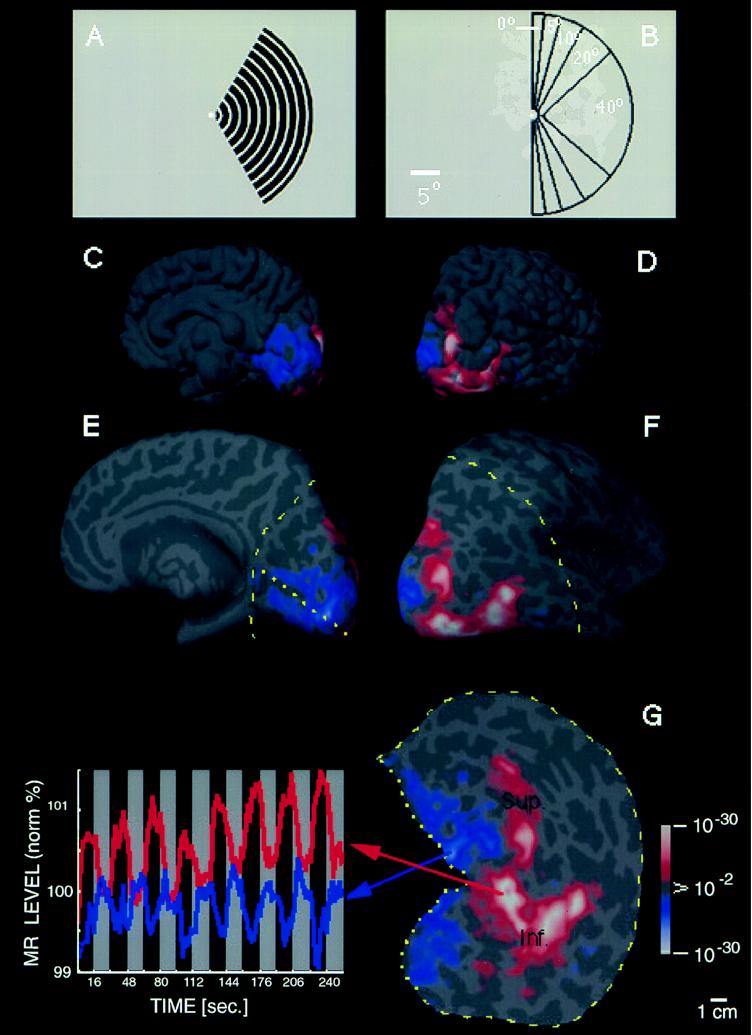
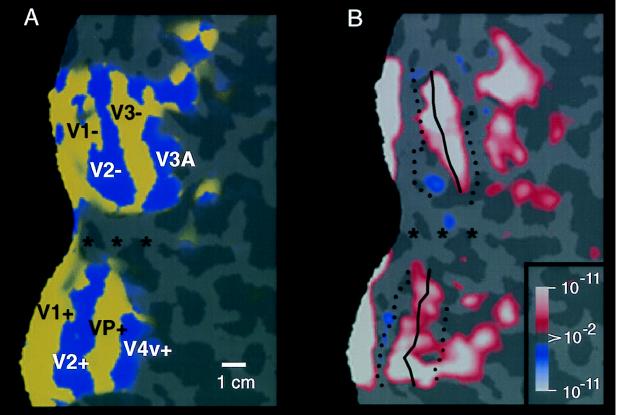
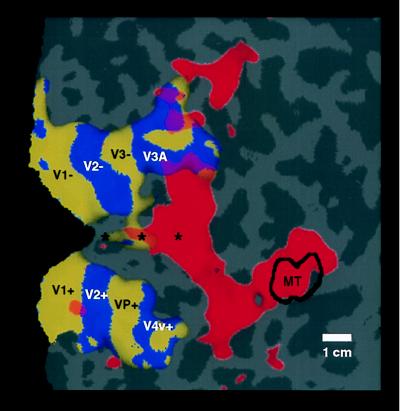
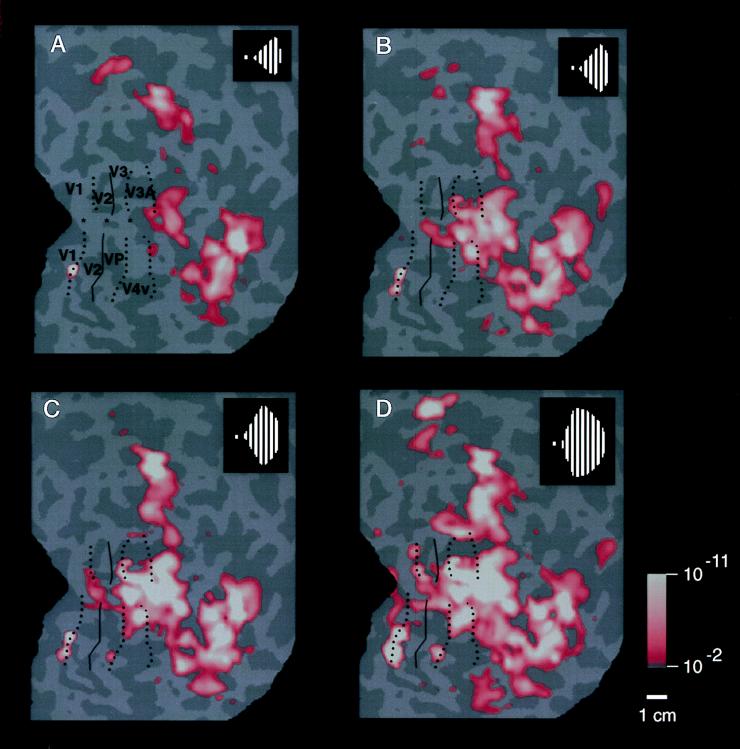
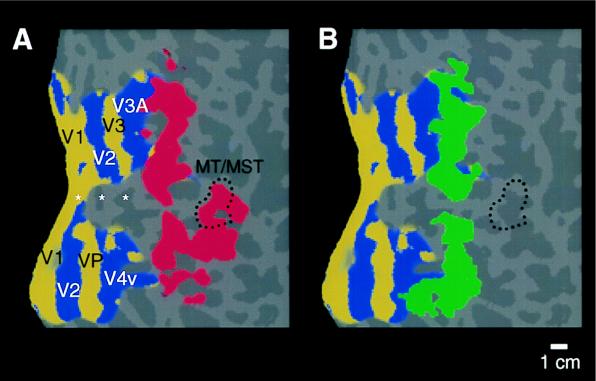
Previous studies of cortical retinotopy focused on influences from the contralateral visual field, because ascending inputs to cortex are known to be crossed. Here, functional magnetic resonance imaging was used to demonstrate and analyze an ipsilateral representation in human visual cortex. Moving stimuli, in a range of ipsilateral visual field locations, revealed activity: (i) along the vertical meridian in retinotopic (presumably lower-tier) areas; and (ii) in two large branches anterior to that, in presumptive higher-tier areas. One branch shares the anterior vertical meridian representation in human V3A, extending superiorly toward parietal cortex. The second branch runs antero-posteriorly along lateral visual cortex, overlying motion-selective area MT. Ipsilateral stimuli sparing the region around the vertical meridian representation also produced signal reductions (perhaps reflecting neural inhibition) in areas showing contralaterally driven retinotopy. Systematic sampling across a range of ipsilateral visual field extents revealed significant increases in ipsilateral activation in V3A and V4v, compared with immediately posterior areas V3 and VP. Finally, comparisons between ipsilateral stimuli of different types but equal retinotopic extent showed clear stimulus specificity, consistent with earlier suggestions of a functional segregation of motion vs. form processing in parietal vs. temporal cortex, respectively.
Figures
Similar articles
-
Second and third visual areas of the cat: interindividual variability in retinotopic arrangement and cortical location.J Physiol. 1980 Feb;299:247-76. doi: 10.1113/jphysiol.1980.sp013123. J Physiol. 1980. PMID: 7381768 Free PMC article.
-
Functional analysis of V3A and related areas in human visual cortex.J Neurosci. 1997 Sep 15;17(18):7060-78. doi: 10.1523/JNEUROSCI.17-18-07060.1997. J Neurosci. 1997. PMID: 9278542 Free PMC article.
-
The retinotopic organization of primate dorsal V4 and surrounding areas: A functional magnetic resonance imaging study in awake monkeys.J Neurosci. 2003 Aug 13;23(19):7395-406. doi: 10.1523/JNEUROSCI.23-19-07395.2003. J Neurosci. 2003. PMID: 12917375 Free PMC article.
-
Clarifying homologies in the mammalian cerebral cortex: the case of the third visual area (V3).Clin Exp Pharmacol Physiol. 2005 May-Jun;32(5-6):327-39. doi: 10.1111/j.1440-1681.2005.04192.x. Clin Exp Pharmacol Physiol. 2005. PMID: 15854138 Review.
-
Cortical connections of MT in four species of primates: areal, modular, and retinotopic patterns.Vis Neurosci. 1990 Aug;5(2):165-204. doi: 10.1017/s0952523800000213. Vis Neurosci. 1990. PMID: 2278944 Review.
Cited by
-
On the role of suppression in spatial attention: evidence from negative BOLD in human subcortical and cortical structures.J Neurosci. 2014 Jul 30;34(31):10347-60. doi: 10.1523/JNEUROSCI.0164-14.2014. J Neurosci. 2014. PMID: 25080595 Free PMC article.
-
Visual speech perception in foveal and extrafoveal vision: further implications for divisions in hemispheric projections.PLoS One. 2014 Jul 17;9(7):e98273. doi: 10.1371/journal.pone.0098273. eCollection 2014. PLoS One. 2014. PMID: 25032950 Free PMC article.
-
Cholinergic enhancement reduces spatial spread of visual responses in human early visual cortex.Neuron. 2008 Dec 10;60(5):904-14. doi: 10.1016/j.neuron.2008.09.038. Neuron. 2008. PMID: 19081383 Free PMC article. Clinical Trial.
-
Multimodal functional neuroimaging: integrating functional MRI and EEG/MEG.IEEE Rev Biomed Eng. 2008;1(2008):23-40. doi: 10.1109/RBME.2008.2008233. Epub 2008 Nov 5. IEEE Rev Biomed Eng. 2008. PMID: 20634915 Free PMC article. Review.
-
Extrastriate visual cortex reorganizes despite sequential bilateral occipital stroke: implications for vision recovery.Front Hum Neurosci. 2015 Apr 28;9:224. doi: 10.3389/fnhum.2015.00224. eCollection 2015. Front Hum Neurosci. 2015. PMID: 25972800 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
