

Shifts in the life history of parasitic wasps correlate with pronounced alterations in early development
- PMID: 9448291
- PMCID: PMC18685
- DOI: 10.1073/pnas.95.3.1097
Shifts in the life history of parasitic wasps correlate with pronounced alterations in early development
Abstract
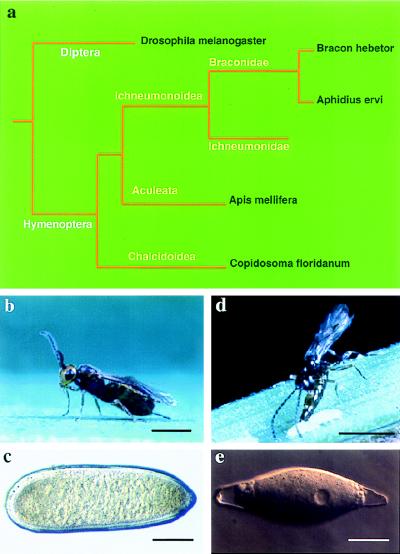
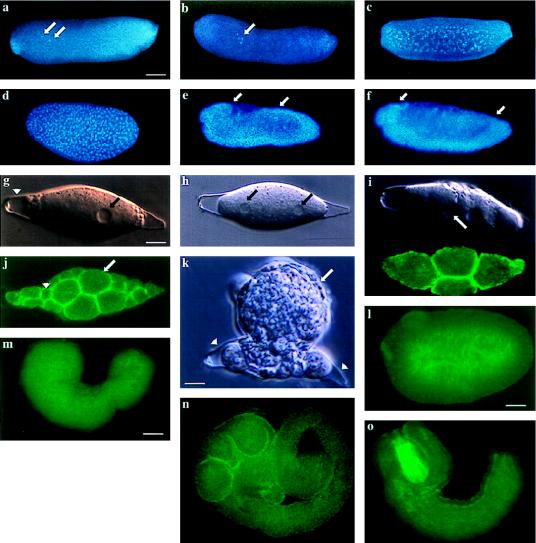
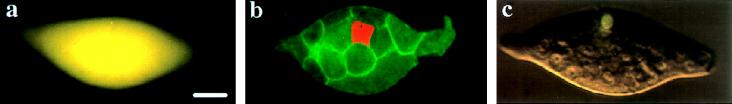
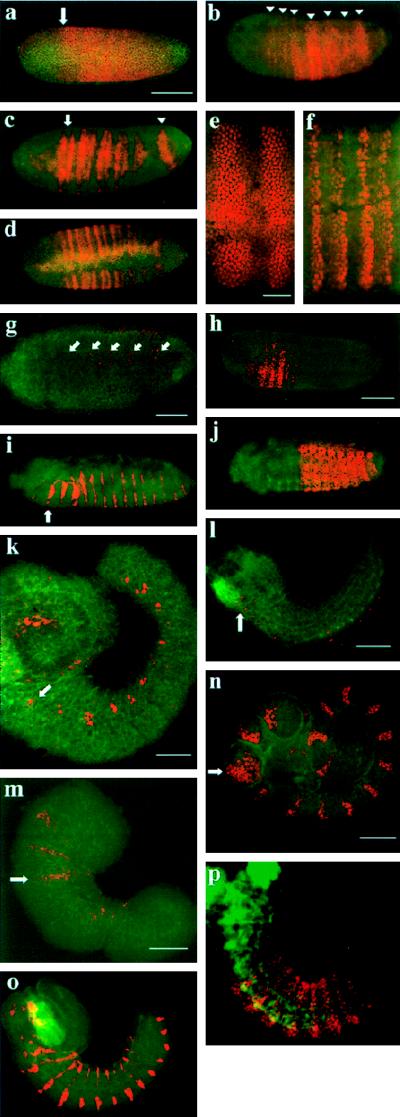
Developmental processes have been traditionally viewed to be invariant within higher taxa. However, examples are known whereby closely related species exhibit alterations in early embryogenesis yet appear very similar as adults. Such developmental changes are thought to occur in response to shifts in life history. In insects, the regulation of embryonic development has been intensively studied in model species like Drosophila melanogaster. Previous comparative studies suggest that the developmental processes documented in Drosophila well describe embryogenesis of advanced, holometabolous, insects generally. There have been few attempts, however, to take into account how life history has influenced early development of insects or to characterize early development of species with life histories fundamentally different from flies. Here we compared early development of two species from the same family of parasitic wasps that exhibit very different life histories. Bracon hebetor is an ectoparasite that lays large, yolky eggs on the integument of its host that develop much like the free-living honeybee and Drosophila. In contrast, Aphidius ervi is an endoparasite that lays small and apparently yolk-free eggs that develop in the hemocoel of the host. This wasp exhibits a radically different mode of early development at both the cellular and molecular level from B. hebetor. The developmental changes in A. ervi reflect functional adaptations for its derived life history and argue that departures from the fly paradigm may occur commonly among insects whose eggs develop under conditions different from typical terrestrial species.
Figures
References
-
- von Baer K E. Entwicklungsgeschichte der Thiere: Beobachtung und Reflexion. Konigsberg, Germany: Born-trager; 1828.
-
- Gould S J. Ontogeny and Phylogeny. Cambridge, MA: Belknap; 1977.
-
- Wray G A. Science. 1995;267:115–116. - PubMed
-
- del Pino E M, Ellinson R P. Nature (London) 1983;306:589–591.
-
- Scott L B, Lennarz W J, Raff R A, Wray G A. Dev Biol. 1990;138:188–193. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
